Trait-Based vs. Directed Evolution: A Strategic Guide for Optimizing Biological Communities and Bioprocesses
This article provides a comprehensive comparison for researchers and drug development professionals between two powerful paradigms for biological optimization: trait-based ecology and directed evolution.
Trait-Based vs. Directed Evolution: A Strategic Guide for Optimizing Biological Communities and Bioprocesses
Abstract
This article provides a comprehensive comparison for researchers and drug development professionals between two powerful paradigms for biological optimization: trait-based ecology and directed evolution. We explore the foundational principles of both approaches, from the analysis of functional traits that govern community performance to the laboratory-driven iterative cycles of genetic diversification and selection. The scope covers modern methodologies, including CRISPR-based diversification and high-throughput screening, alongside practical strategies for troubleshooting yield variation and process control. By synthesizing insights from theoretical ecology and applied biotechnology, this guide offers a validated framework for selecting the optimal strategy to engineer microbes, cellular communities, and biocatalysts for enhanced robustness, yield, and novel function in biomedical applications.
Core Principles: From Natural Trait Diversity to Laboratory-Driven Evolution
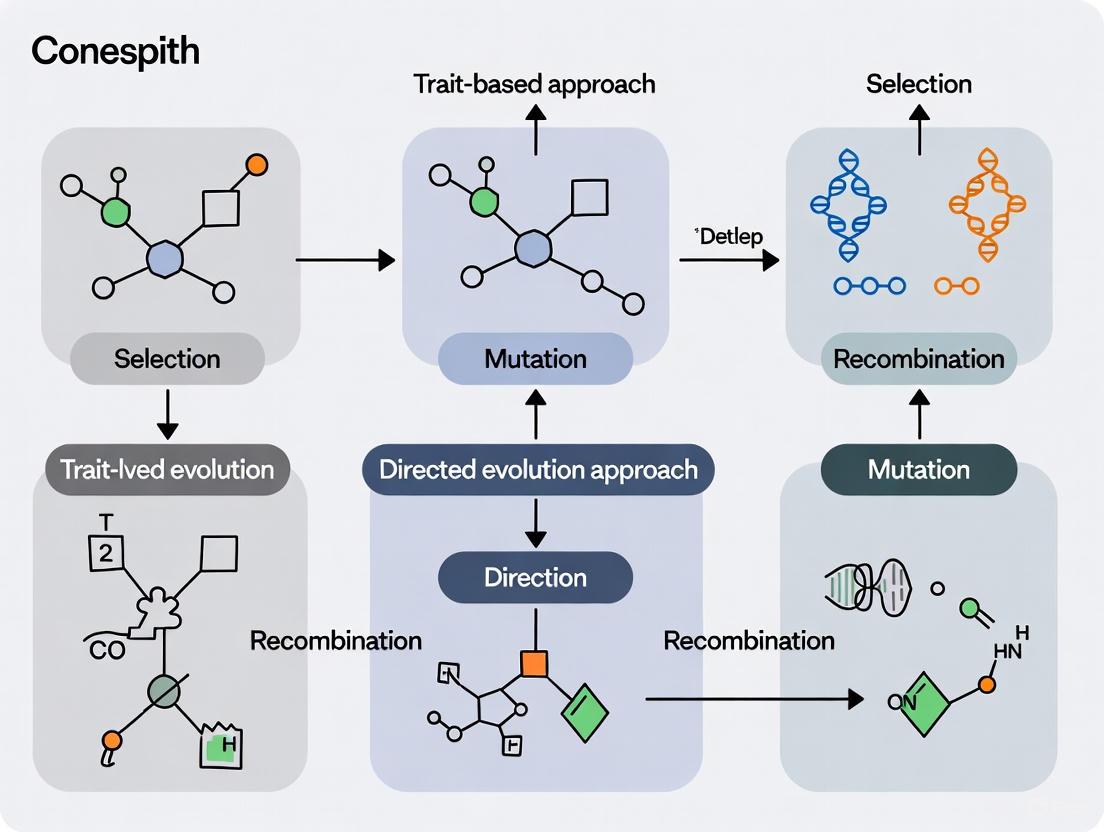
In the pursuit of optimizing biological systems for research and therapeutic applications, two powerful methodologies have emerged: trait-based evolution and directed evolution. While both approaches harness evolutionary principles, they differ fundamentally in their philosophy, implementation, and application. Trait-based evolution is an analytical framework rooted in quantitative genetics that investigates how phenotypic traits evolve in natural populations or communities in response to selective pressures. In contrast, directed evolution is an engineering methodology that mimics natural selection in laboratory settings to optimize biomolecules for specific human-defined functions. This guide provides a comprehensive comparison of these paradigms, offering researchers a clear framework for selecting appropriate strategies for biological optimization challenges.
Core Principles and Theoretical Foundations
Trait-Based Evolution
Trait-based evolution approaches investigate how heritable phenotypic characteristics change in populations over time in response to environmental selective pressures [1]. This framework is grounded in quantitative genetics, which analyzes the evolution of continuous traits influenced by multiple genes and environmental factors. The core mathematical framework is described by the Lande equation: Δz̄ = G∇ȳ, where Δz̄ represents the change in mean phenotype, G is the genetic variance-covariance matrix, and ∇ȳ is the selection gradient [1]. This equation highlights that evolutionary response depends not only on the strength of selection but also on the available genetic variation and its structure.
These approaches typically study evolution in natural populations or controlled experimental settings where selective pressures are observed or manipulated rather than designed. Research focuses on understanding fundamental evolutionary processes, including how traits evolve in response to environmental changes, species interactions, and community dynamics [1] [2]. A key consideration is the evolution of trait variance itself—as species diversity increases in communities, competition often drives the evolution of narrower trait breadths in individual species, which can surprisingly reduce overall functional diversity despite increased species richness [2].
Directed Evolution
Directed evolution is a protein engineering methodology that mimics natural evolution in laboratory settings to optimize biomolecules for specific applications [3] [4]. This approach involves iterative cycles of genetic diversification and selection to rapidly isolate improved variants without requiring detailed mechanistic understanding of the sequence-function relationship [5] [6].
The fundamental process consists of creating genetic diversity in a target gene through random or targeted mutagenesis, expressing these variants to create a library, screening or selecting for desired properties, and using improved variants as templates for subsequent evolution cycles [4]. Unlike natural evolution, directed evolution operates on human-defined objectives and typically occurs over much shorter timescales—weeks or months rather than centuries [3]. This methodology effectively navigates complex fitness landscapes by exploring sequence spaces that would be difficult to predict rationally, making it particularly valuable for engineering proteins with novel functions or improved stability [5] [4].
Table 1: Fundamental Characteristics of Trait-Based and Directed Evolution Approaches
| Characteristic | Trait-Based Evolution | Directed Evolution |
|---|---|---|
| Primary Objective | Understand natural evolutionary processes | Engineer biomolecules with desired properties |
| Theoretical Basis | Quantitative genetics, population genetics | Molecular evolution, protein engineering |
| Typical Context | Natural populations, ecological communities | Laboratory experiments, industrial applications |
| Timescale | Generational (medium to long-term) | Rapid cycles (days to weeks) |
| Genetic Diversity Source | Standing variation, new mutations | Artificially generated mutations |
| Key Outcome | Understanding of evolutionary patterns | Optimized biomolecules for specific applications |
Methodologies and Experimental Protocols
Trait-Based Evolution Workflows
Trait-based evolution research employs both observational and experimental approaches to investigate evolutionary processes. Long-term observational studies monitor natural populations over extended periods, such as the landmark research on Darwin's finches that has documented evolutionary changes in beak size and shape in response to climatic variations over four decades [7]. These studies incorporate natural environmental complexity, population demographics, and species interactions without experimental manipulation.
Experimental field approaches manipulate selective pressures in natural settings to establish causal relationships between environmental factors and evolutionary outcomes. Examples include long-term studies of guppies in Trinidadian streams, where researchers introduced predators to different populations and observed subsequent evolutionary changes in life history traits [7]. Laboratory selection experiments provide greater environmental control, enabling researchers to examine evolutionary dynamics across thousands of generations while maintaining replicate populations. The Long-Term Evolution Experiment (LTEE) with Escherichia coli, ongoing for over 75,000 generations, has revealed fundamental principles about adaptive evolution, historical contingency, and the dynamics of trait evolution [7].
Directed Evolution Workflows
Directed evolution employs systematic laboratory protocols to engineer biomolecules through iterative diversification and selection. The following workflow diagram illustrates a generalized directed evolution pipeline:
Library Generation Methods create genetic diversity through various mutagenesis strategies. Error-prone PCR introduces random point mutations throughout the gene sequence using reaction conditions that reduce polymerase fidelity [3] [4]. DNA shuffling recombines fragments from homologous genes to create chimeric proteins, mimicking natural recombination [4]. Site-saturation mutagenesis targets specific residues to explore all possible amino acid substitutions at chosen positions [3].
Selection and Screening Strategies enable identification of improved variants. Display techniques (phage, yeast, or ribosome display) physically link genotype to phenotype, allowing high-throughput screening of binding affinity or other properties [3]. Fluorescence-activated cell sorting (FACS) enables ultra-high-throughput screening of cellular properties when the desired function can be linked to fluorescence [3] [5]. Microtiter plate assays facilitate medium-throughput screening of enzymatic activity using colorimetric or fluorimetric substrates [3].
Recent methodological advances focus on optimizing selection conditions to maximize efficiency. Approaches include Design of Experiments (DoE) to systematically evaluate multiple selection parameters and deep sequencing of selection outputs to identify significantly enriched mutants at appropriate coverage levels [5].
Table 2: Key Methodological Techniques in Directed Evolution
| Method Category | Specific Techniques | Key Applications | Throughput |
|---|---|---|---|
| Library Generation | Error-prone PCR, DNA shuffling, Site-saturation mutagenesis, RAISE, TRINS | Creating genetic diversity in target genes | Varies by technique |
| Selection/Screening | Phage display, FACS, Microplate assays, QUEST, Cofactor regeneration coupling | Identifying variants with desired properties | 10^3 - 10^13 variants |
| Analysis | Next-generation sequencing, Functional characterization, Kinetic analysis | Validating improved variants and understanding mutations | Dependent on variant number |
Applications in Research and Drug Development
Trait-Based Evolution Applications
Trait-based approaches provide critical insights for evolutionary biology, conservation, and understanding disease dynamics. In evolutionary rescue research, quantitative genetic models investigate whether populations can adapt rapidly enough to avoid extinction in changing environments, with implications for conservation biology amid climate change [1]. Cancer evolution studies apply evolutionary models to understand how tumor cell populations evolve in response to treatment, investigating how birth-death rate tradeoffs and cellular turnover influence evolutionary trajectories and therapy resistance [8].
Community ecology studies examine how trait evolution shapes species interactions and ecosystem functioning, revealing that increased species diversity does not necessarily increase functional diversity due to competitive constraints on trait variances [2]. Long-term evolutionary studies connect microevolutionary processes to macroevolutionary patterns, addressing fundamental questions about speciation, evolutionary innovations, and the emergence of complex traits [7].
Directed Evolution Applications
Directed evolution has revolutionized protein engineering for therapeutic and industrial applications. Enzyme engineering creates optimized biocatalysts with enhanced stability, activity, or novel substrate specificity for industrial processes and synthetic biology [3] [4]. Therapeutic protein engineering generates improved antibodies, hormones, and other biologics with enhanced potency, stability, or reduced immunogenicity [3].
Xenobiotic nucleic acid (XNA) polymerase engineering develops specialized enzymes capable of synthesizing and reverse-transcribing artificial genetic polymers with applications in therapeutics and biotechnology [5]. Transcription factor engineering rewires regulatory proteins to respond to novel inducters or regulate new DNA sequences, expanding the synthetic biology toolbox for genetic circuit construction [6]. Metabolic pathway engineering optimizes multi-enzyme pathways for production of pharmaceuticals, biofuels, and valuable chemicals through iterative optimization of pathway components [6] [4].
Research Reagent Solutions Toolkit
Table 3: Essential Research Reagents for Evolution Approaches
| Reagent Category | Specific Examples | Function in Research |
|---|---|---|
| Mutagenesis Systems | Error-prone PCR kits, Chemical mutagens, Transposon-based mutagenesis systems | Introducing genetic diversity for directed evolution or experimental evolution studies |
| Display Technologies | Phage display systems, Yeast surface display, Ribosome display | High-throughput screening of protein-ligand interactions |
| Selection Tools | FACS systems, Microplate readers, Selective growth media | Identifying variants with desired properties from libraries |
| Analysis Reagents | Next-generation sequencing kits, Antibodies for detection, Enzyme substrates | Characterizing evolved variants and their properties |
| Specialized Polymerases | XNA polymerases, Error-prone polymerases, High-fidelity PCR enzymes | Enzymatic tools for library construction and specialized applications |
| (E)-Hex-3-en-1-ol-d2 | (E)-Hex-3-en-1-ol-d2, MF:C6H12O, MW:102.17 g/mol | Chemical Reagent |
| (S)-Sabutoclax | (S)-Sabutoclax, MF:C42H42N2O8S, MW:734.9 g/mol | Chemical Reagent |
Comparative Analysis and Strategic Implementation
Methodological Synergies
While trait-based and directed evolution approaches differ in implementation, they offer complementary insights. Trait-based models can inform directed evolution strategies by predicting how traits might evolve under specific selective pressures [1] [2]. Conversely, directed evolution experiments provide empirical tests of evolutionary hypotheses generated by trait-based theories [4]. This synergy is particularly valuable for understanding complex evolutionary phenomena such as epistasis, historical contingency, and the emergence of novel functions.
Selection Guidelines
Researchers should consider several factors when choosing between these approaches:
Opt for trait-based evolution when:
- Studying natural evolutionary processes in ecological or population contexts
- Investigating evolutionary responses to environmental changes
- Understanding the genetic architecture of complex traits
- Modeling long-term evolutionary dynamics
Choose directed evolution when:
- Engineering biomolecules with specific, human-defined properties
- Optimizing proteins, pathways, or genetic circuits for industrial or therapeutic applications
- Exploring sequence-function relationships without detailed structural knowledge
- Generating novel biocatalysts not found in nature
Consider hybrid approaches when:
- Engineering complex traits influenced by multiple genes
- Developing evolutionary-informed biomolecule engineering strategies
- Testing evolutionary hypotheses through synthetic biology approaches
Trait-based and directed evolution represent distinct yet complementary paradigms for investigating and harnessing evolutionary processes. Trait-based approaches provide the theoretical foundation and analytical tools for understanding how traits evolve in natural systems, while directed evolution offers powerful engineering methodologies for optimizing biomolecules to address human needs. As both fields advance, integrating their insights and methodologies promises to accelerate progress in fundamental evolutionary biology, drug development, and biotechnology. Researchers can leverage the comparative frameworks presented here to select appropriate strategies for their specific biological optimization challenges.
The Theoretical Basis of Trait-Based Ecology and Community Assembly
The quest to understand and engineer biological communities has coalesced around two distinct philosophical and methodological approaches: trait-based ecology and directed evolution. Trait-based ecology represents a rational, design-forward framework where communities are constructed or analyzed based on functional characteristics of their constituent organisms [9] [10]. In contrast, directed evolution embraces a selection-based paradigm, applying iterative cycles of diversification and artificial selection to steer microbial consortia toward desired functional outcomes without requiring detailed knowledge of underlying mechanisms [11]. This guide provides a comparative analysis of these approaches, examining their theoretical foundations, methodological implementations, and performance across research applications, with particular relevance for scientific and drug development professionals seeking to optimize microbial communities for therapeutic or industrial applications.
Theoretical Foundations and Mechanisms
Core Principles of Trait-Based Ecology
Trait-based ecology operates on the fundamental premise that measurable organismal characteristics (traits) directly influence ecological performance and ecosystem functioning [12] [10]. This approach defines functional traits as morpho-physio-phenological characteristics that directly or indirectly link to fitness, growth, reproduction, and survival [13] [10]. The theoretical framework posits that community assembly is governed by environmental filtering—where abiotic conditions select for species possessing traits suitable for that environment—and biotic interactions, including competition, facilitation, and niche differentiation [14] [13].
A key theoretical advancement in trait-based ecology is the recognition that the relationship between species diversity and functional diversity is not necessarily positive. Research on Galápagos land snails demonstrated that in species-rich communities, competition can drive trait narrowing as species evolve narrower trait breadths to avoid competition, potentially reducing overall functional diversity [2]. This challenges the conventional assumption that increasing species diversity automatically enhances ecosystem functionality.
Trait-based approaches further theorize that environmental gradients filter species according to their traits, leading to predictable community assembly patterns. Studies across subtropical karst forests reveal consistent trait-mediated patterns where deciduous forests in drier, fertile soils exhibit resource acquisition traits (e.g., high specific leaf area), while evergreen forests in moist, infertile conditions display resource conservation traits (e.g., high leaf dry matter content) [14].
Core Principles of Directed Evolution
Directed evolution of microbial communities (DEMC) applies artificial selection principles to steer community-level functions through iterative diversification and selection cycles [11]. Unlike trait-based approaches that require deep understanding of functional mechanisms, DEMC operates agnostically to underlying interactions, instead harnessing selective pressures to enrich communities with desired functionalities [11].
The theoretical foundation of DEMC rests on several key principles: (1) microbial communities can be treated as selectable units, (2) community-level functions respond to selective pressures through compositional and functional shifts, and (3) iterative selection stabilizes desirable functional attributes. This approach leverages natural evolutionary processes—mutation, horizontal gene transfer, and selection—but directs them toward specific functional goals [11] [15].
Research in fermented food systems demonstrates that DEMC can enhance functional stability and performance without genetic engineering. For instance, microbial consortia subjected to acid stress develop improved acid resistance and metabolic performance under extreme pH conditions, illustrating how ecological adaptation through directed evolution produces robust functional outcomes [11].
Comparative Theoretical Framework
Table 1: Theoretical Comparison Between Trait-Based and Directed Evolution Approaches
| Theoretical Aspect | Trait-Based Ecology | Directed Evolution |
|---|---|---|
| Fundamental Principle | Environmental filtering and niche-based assembly | Artificial selection and evolutionary steering |
| Knowledge Requirement | Requires prior trait-function knowledge | Agnostic to mechanistic understanding |
| Timescale | Primarily ecological (contemporary) | Eco-evolutionary (multiple generations) |
| Unit of Selection | Individual traits/species | Whole community as selectable unit |
| Predictability | High when trait-function relationships known | Emergent through selection history |
| Stability Mechanisms | Niche differentiation and complementarity | Selected functional redundancy |
Methodological Implementation
Trait-Based Experimental Workflows
Trait-based methodologies follow rational design principles beginning with trait characterization and culminating in community assembly or analysis. The experimental workflow typically involves: (1) identifying relevant functional traits for target ecosystem functions, (2) measuring trait values across candidate species, (3) constructing communities based on trait compatibility and complementarity, and (4) validating functional outcomes [9] [16].
Advanced trait-based approaches incorporate phylogenetic inference to predict trait values when direct measurements are unavailable. Research on infant gut microbiome development employed a phylogeny-based method using 16S rRNA data and curated trait databases to infer traits like oxygen tolerance, sporulation ability, and 16S rRNA gene copy number across microbial taxa [13]. This approach enables trait-based analysis even when comprehensive trait measurements are lacking.
Table 2: Key Methodological Components in Trait-Based Ecology
| Methodological Component | Description | Application Example |
|---|---|---|
| Community-Weighted Means (CWM) | Mean trait values weighted by species abundance | Quantifying dominant traits in karst forests [14] |
| Trait-Based Null Models | Statistical tests comparing observed trait distributions to random expectations | Identifying environmental filtering in plant communities [14] |
| Phylogenetic Trait Imputation | Using evolutionary relationships to infer unknown traits | Predicting microbial traits in infant gut development [13] |
| Functional Diversity Metrics | Quantifying the volume of trait space occupied by a community | Relating species diversity to functional diversity [2] |
Directed Evolution Methodologies
Directed evolution implements iterative selection cycles consisting of four key phases: (1) initial community construction, (2) application of selective pressure, (3) functional screening, and (4) propagation of superior communities [11] [15]. This design-build-test-learn cycle continues until communities stably maintain target functions.
In fermented food applications, DEMC typically begins with diverse starter communities from spontaneous fermentation, backslopping, or defined starters [11]. Selective pressures are then applied based on target functionalities—temperature stress for thermostability, acid stress for pH tolerance, or substrate limitations for metabolic efficiency. Functional screening identifies communities exhibiting enhanced performance, which are then propagated for subsequent selection cycles.
Recent technological innovations have enhanced DEMC capabilities. The PROTEUS platform uses chimeric virus-like vesicles to enable directed evolution in mammalian cells, addressing previous limitations in mammalian directed evolution systems [17]. This platform demonstrates how viral vectors can maintain system integrity across extended evolution campaigns while generating sufficient diversity for functional optimization.
Experimental Protocols for Community Optimization
Protocol 1: Trait-Based Community Assembly
- Initial Characterization: Quantify key functional traits across candidate species pool (e.g., specific leaf area, wood density for plants; oxygen tolerance, substrate utilization for microbes) [14] [13]
- Trait-Function Modeling: Establish statistical relationships between trait values and target functions using regression models or machine learning
- Community Design: Select species combinations that optimize trait complementarity based on functional objectives
- Validation: Assemble synthetic communities and measure functional outcomes versus predictions
- Refinement: Adjust trait-function models based on empirical results
Protocol 2: Directed Evolution of Microbial Communities
- Community Initiation: Construct initial diverse community from environmental samples, culture collections, or defined isolates [11]
- Diversification Phase: Allow natural mutation, horizontal gene transfer, or introduce random mutagenesis to generate variation
- Selection Pressure: Apply targeted stressor relevant to desired function (e.g., high temperature, acidic pH, antibiotic exposure)
- Screening and Propagation: Identify high-performing communities through functional assays; transfer superior communities to fresh medium
- Iteration: Conduct multiple cycles (typically 5-20) of diversification and selection
- Stabilization: Monitor functional stability across transfers without selection pressure
Performance Comparison and Experimental Data
Functional Optimization Efficiency
Table 3: Comparative Performance of Trait-Based versus Directed Evolution Approaches
| Performance Metric | Trait-Based Ecology | Directed Evolution | Experimental Evidence |
|---|---|---|---|
| Time to Optimization | Weeks to months | Months to multiple generations | DEMC requires 5-20 cycles for fermented foods [11] |
| Functional Precision | High for known traits | Emergent, potentially superior for complex functions | Trait-based approaches enable precise division of labor [9] |
| Stability Outcomes | Variable, requires careful design | Often high through selected adaptations | DEMC communities maintain function under stress [11] |
| Novel Function Discovery | Limited to designed combinations | Can yield unexpected solutions | DEMC produces novel regulatory tools [17] |
| Knowledge Dependency | High prior knowledge required | Minimal prior knowledge needed | Trait-based relies on established trait-function relationships [10] |
| Technical Barriers | Trait measurement, modeling | High-throughput screening | DEMC benefits from automated screening [11] |
Applications in Biotechnology and Medicine
Trait-based approaches have demonstrated particular success in environmental biotechnology where trait-function relationships are well-characterized. Synthetic microbial communities (SynComs) designed using trait-based principles show enhanced performance in pollutant degradation, where complementary metabolic traits are strategically combined [16]. Similarly, agricultural applications employ trait-based design to assemble plant growth-promoting rhizobacteria consortia with balanced competitive and cooperative interactions [16].
Directed evolution excels in optimizing complex, multifunctional outcomes where mechanistic understanding is limited. In fermented food production, DEMC has improved both sensory qualities and production efficiency simultaneously [11]. Medical applications include evolving microbial communities for gut microbiome therapeutics, where DEMC can steer communities toward stable, beneficial configurations without requiring complete understanding of underlying microbial interactions.
Emerging platforms like PROTEUS enable directed evolution of protein functions within mammalian cells, generating tools with mammalian-specific adaptations [17]. This technology has evolved tetracycline-controlled transactivators with altered doxycycline responsiveness, creating more sensitive genetic regulation tools for biotechnology and therapeutic applications.
Research Reagent Solutions
Table 4: Essential Research Tools for Community Optimization Studies
| Research Tool | Function | Application Context |
|---|---|---|
| PROTEUS Platform | Mammalian directed evolution using virus-like vesicles | Protein evolution in mammalian cellular environment [17] |
| Error-Prone PCR | In vitro random mutagenesis | Generating diverse gene variants for directed evolution [15] |
| Multi-Omics Analysis | Comprehensive community characterization | Tracking compositional and functional changes in both approaches [16] [11] |
| Trait Databases | Curated functional trait data | Informing trait-based design across taxa [13] [10] |
| CRISPR-Cas Systems | Targeted genome editing | Creating specific variants in trait-based design [15] |
| Automated Cultivation Systems | High-throughput community propagation | Scaling directed evolution experiments [11] |
| Genome-Scale Metabolic Models | Predicting metabolic interactions | Informing trait-based community design [16] |
Trait-based ecology and directed evolution represent complementary rather than competing paradigms for community optimization. Trait-based approaches offer precision and predictability when mechanistic understanding is sufficient, while directed evolution provides powerful functionality discovery when systems are too complex for rational design. The most effective community engineering strategies will likely integrate both approaches—using trait-based principles to design initial communities and guide selective pressures, while employing directed evolution to refine and stabilize functional outcomes. This synergistic framework promises to accelerate development of microbial consortia for therapeutic applications, bioproduction, and environmental restoration, ultimately enhancing our ability to harness biological communities for human needs.
Directed evolution stands as one of the most powerful tools in protein engineering, mimicking the principles of natural selection to steer biological molecules toward user-defined goals. This method has transformed basic biological research and the development of therapeutics, enabling engineers to create proteins with enhanced stability, novel catalytic activities, and specific binding affinities without requiring prior structural knowledge. This guide traces the pivotal milestones in the history of directed evolution, objectively compares the performance of its key methodologies, and situates these advances within the broader thesis of trait-based versus directed evolution approaches for community optimization research. For the researcher, understanding this evolution is critical for selecting the optimal strategy for a given protein engineering challenge.
Historical Timeline: Key Experiments and Methodologies
The following table chronicles the major developments in directed evolution, from foundational concepts to contemporary, computationally-enhanced practices.
Table 1: Historical Milestones in Directed Evolution
| Year(s) | Milestone / Experiment | Key Methodology | Impact & Performance Outcome |
|---|---|---|---|
| 1960s | Spiegelman's Monster [18] [19] | In vitro evolution of a self-replicating RNA molecule using Qβ replicase, selecting for fastest-replicating variants. | Proof-of-concept: Demonstrated that molecules could be evolved artificially under selective pressure, resulting in a minimal, highly efficient 218-nucleotide RNA [19]. |
| 1980s | Development of Phage Display [18] [4] | Selection: A library of peptide variants is displayed on the surface of bacteriophages; binding to a target antigen enables isolation of high-affinity binders. | Enabled affinity maturation: Revolutionized antibody and peptide engineering. A direct lineage led to modern therapeutic antibodies [18]. |
| 1990s | Modern Directed Evolution of Enzymes [18] [4] | Random Mutagenesis & Screening: Iterative rounds of error-prone PCR and high-throughput screening for improved traits (e.g., stability, activity). | Established the modern paradigm: A seminal study evolved subtilisin E, achieving a 256-fold increase in activity in a non-aqueous solvent (DMF) [4]. |
| 1994 | DNA Shuffling [4] | In vitro Homologous Recombination: DNA fragments from a gene family are randomly reassembled to create chimeric libraries. | Accelerated evolution: Mimicked sexual recombination. Evolution of a β-lactamase increased host antibiotic resistance by 32,000-fold, far surpassing non-recombinogenic methods [4]. |
| 2018 | Nobel Prize in Chemistry [18] | Collective recognition of Frances Arnold (directed evolution of enzymes), and George Smith and Gregory Winter (phage display). | Field validation: Cemented the transformative impact of directed evolution across basic science and medicine, particularly for generating therapeutic antibodies and green industrial catalysts. |
| Present / Future | Integration with AI & Machine Learning [20] [21] [22] | Semi-rational Design: Machine learning models predict fitness landscapes from sequence-activity data, guiding library design and identifying beneficial mutations. | Enhanced efficiency: Reduces experimental burden by focusing on promising sequence regions. AI tools like AlphaFold (for structure prediction) and RFdiffusion (for de novo design) are creating powerful synergies with directed evolution [21] [22]. |
Experimental Protocols in Directed Evolution
The core process of directed evolution is an iterative cycle, but the specific protocols for library generation and selection have diversified significantly.
The General Directed Evolution Workflow
The universal process involves repeated rounds of three steps: Diversification, Selection (or Screening), and Amplification [18]. The workflow is illustrated below.
Detailed Methodologies for Library Creation and Selection
Table 2: Key Experimental Protocols in Directed Evolution
| Method Category | Specific Protocol | Detailed Methodology | Typical Library Size | Best Use Case |
|---|---|---|---|---|
| Random Mutagenesis | Error-Prone PCR [18] [4] | PCR is performed under conditions that reduce fidelity (e.g., unbalanced dNTPs, Mn2+ ions), introducing random point mutations throughout the gene. | 10^4 - 10^6 variants | Broad exploration of local sequence space around a parent sequence. |
| Recombination | DNA Shuffling [4] | A family of homologous genes is digested with DNase I, and the fragments are reassembled in a primer-free PCR, creating chimeric genes. | 10^6 - 10^12 variants | Exploring diversity from natural homologs or beneficial mutants from earlier rounds. |
| Selection | Phage Display [18] | A library of protein variants is displayed on the surface of phage particles. Phages binding to an immobilized target are retained and eluted, and their DNA is amplified. | Up to 10^11 variants | Isolating high-affinity binders (e.g., antibodies, peptides). |
| Screening | Fluorescence-Activated Cell Sorting (FACS) [20] | Cells displaying or expressing protein variants are individually analyzed for a fluorescent signal linked to activity. The top-performing fraction is isolated and sorted. | 10^7 - 10^9 variants per hour | Quantifying and isolating variants based on activity levels when a fluorescent reporter is available. |
| Targeted Mutagenesis | Focused Libraries [18] | Based on structural knowledge or computational predictions, specific residues (e.g., in the active site) are randomized using degenerate codons. | 10^2 - 10^5 variants | Efficiently optimizing a specific region of a protein without the noise of global mutagenesis. |
The Evolutionary Design Spectrum: A Unifying Framework
A modern perspective posits that all biological design processes, from traditional design to directed evolution, exist on a unified evolutionary design spectrum [23]. This framework is defined by two key dimensions: the throughput (number of variants tested per cycle) and the number of generations (design cycles). This spectrum directly informs the trait-based versus directed evolution debate.
Trait-Based (Rational) Design: This approach lies on one end of the spectrum, characterized by low throughput but high prior knowledge exploitation. Engineers use structural and mechanistic information to design specific changes, typically testing a small number of variants in a single, knowledge-driven cycle [18] [23].
Directed Evolution: This approach lies on the other end, characterized by high throughput and high exploration over multiple generations. It requires minimal prior knowledge, relying instead on iterative random variation and selection to discover beneficial traits [18] [23].
Semi-Rational (AI-Guided) Design: This hybrid approach is emerging as a powerful middle ground. It uses machine learning to exploit large datasets (knowledge) to create focused libraries, thereby guiding the exploration process more efficiently. This enhances the power of directed evolution by reducing its reliance on pure randomness and massive screening [20] [21] [22].
The Scientist's Toolkit: Essential Research Reagents
Successful directed evolution experiments rely on a suite of specialized reagents and biological tools.
Table 3: Key Research Reagent Solutions for Directed Evolution
| Reagent / Material | Function in the Workflow |
|---|---|
| Error-Prone PCR Kit | A optimized blend of polymerase and buffer conditions to introduce random mutations during gene amplification [18]. |
| Phage Display Vector | A plasmid engineered for the surface display of peptide/protein fusions on bacteriophages, enabling selection [18]. |
| Fluorescent Substrate/Reporter | A molecule that produces a measurable fluorescent signal upon enzymatic activity, enabling high-throughput screening via FACS or microplate readers [20] [18]. |
| Microfluidic Cell Sorter | Advanced instrumentation that allows for high-throughput, single-cell analysis and sorting based on phenotypic traits, enabling complex selection strategies [20]. |
| In vivo Mutagenesis Strain | Engineered host cells (e.g., E. coli) with hypermutable genomes or inducible mutagenesis systems targeted to a plasmid of interest [20]. |
| Yeast Surface Display System | A eukaryotic platform for displaying protein libraries on the yeast surface, allowing for the selection of well-folded, glycosylated proteins [21]. |
| Loloatin B | Loloatin B|Cyclic Decapeptide Antibiotic|RUO |
| Veldoreotide TFA | Veldoreotide TFA, MF:C62H75F3N12O12, MW:1237.3 g/mol |
The journey of directed evolution, from Spiegelman's minimalist RNA to today's AI-integrated protein engineering platforms, demonstrates a clear trajectory toward increasingly sophisticated and efficient design. The historical comparison reveals that no single methodology is universally superior; rather, the choice between high-exploration directed evolution and high-exploitation trait-based design depends on the available knowledge and the specific engineering goal. The emerging paradigm, however, powerfully synthesizes these approaches. By leveraging computational models to learn from fitness landscapes, modern protein engineers can now design smarter libraries and navigate sequence space with unprecedented precision. This semi-rational, data-driven approach represents the current frontier, optimizing the community of protein variants by intelligently balancing the exploration of new traits with the exploitation of known structural principles.
This guide compares two fundamental approaches in evolutionary optimization: trait-based evolution, which observes how natural communities evolve in response to environmental pressures, and directed evolution, an experimental technique that mimics natural evolution to engineer biomolecules with desired properties. The comparison is framed for researchers aiming to optimize biological systems, from microbial communities to therapeutic proteins.
Core Conceptual Comparison
The table below outlines the foundational principles of each approach.
| Feature | Trait-Based Evolution (Eco-Evolutionary) | Directed Evolution (Protein Engineering) |
|---|---|---|
| Core Principle | Studies how heritable functional traits influence fitness and ecosystem function in natural environments [24]. | Harnesses Darwinian principles in a laboratory setting to rapidly evolve biomolecules with enhanced functions [3]. |
| Primary Context | Natural ecosystems and communities (e.g., plant communities, land snails) [24] [2]. | In vitro or in vivo laboratory systems [3]. |
| Driving Force | Natural selection from environmental pressures (e.g., competition, abiotic stress) [2]. | Artificial selection for human-defined objectives (e.g., enzyme activity, drug binding) [3]. |
| Key Metrics | Specific Effect Function (SEF), Specific Response Function (SRF), functional diversity, phylogenetic signal [24]. | Library size, enrichment factor, catalytic efficiency ((k{cat}/Km)), binding affinity ((K_d)) [3]. |
| Typical Workflow | 1. Trait measurement → 2. Environmental correlation → 3. Fitness consequence → 4. Phylogenetic analysis [24] | 1. Library generation → 2. Selection/Screening → 3. Characterization → 4. Iteration [3] |
Experimental Protocols in Practice
This section details the standard methodologies for both approaches, providing a blueprint for experimental design.
Trait-Based Community Analysis
This protocol is used to understand how communities assemble and function in response to environmental drivers [24] [2].
- Trait Selection and Measurement: Identify and quantify functional traits relevant to the ecosystem process and environmental driver of interest. For plants, this may include leaf nitrogen content (effect trait) and seed mass (response trait). For animals, it could involve body size or feeding structures [24].
- Environmental Grading: Characterize the environmental gradient across study sites (e.g., nutrient availability, temperature, presence of heavy metals or biocides) [25].
- Community Assessment: Census species abundance and distribution across the environmental gradient.
- Data Integration and Modeling:
- Calculate Specific Effect Function (SEF) and Specific Response Function (SRF) for key species [24].
- Analyze the correlation between SEF and SRF and test for phylogenetic signal to assess ecosystem vulnerability [24].
- Use models to test if trait variance decreases in species-rich communities due to competitive packing [2].
Directed Evolution of a Protein
This general workflow is employed to improve or alter enzyme activity, binding affinity, or stability [3].
- Library Generation: Create genetic diversity in a parent gene sequence.
- Selection or Screening: Identify improved variants from the library.
- High-Throughput Selection: Use methods like phage display or FACS to isolate binders or enzymes with desired activity from large libraries (>10^8 members) [3].
- Lower-Throughput Screening: Screen smaller libraries (<10^4 members) using plate-based assays (e.g., colorimetric or fluorimetric changes) [3].
- Characterization: Purify the hit variants and characterize them using relevant biochemical assays (e.g., kinetics, thermostability).
- Iteration: Use the best-performing variant as a template for subsequent rounds of diversification and selection to cumulatively improve the function.
Experimental Data and Comparison
The following table synthesizes quantitative data and outcomes from studies employing these approaches.
| Approach / Study | Key Experimental Data | Outcome / Implication |
|---|---|---|
| Trait-Based: Land Snails (Galápagos) [2] | In communities with higher species diversity, individual species evolved narrower trait breadths (reduced intraspecific variance). | Increased species diversity led to reduced functional diversity due to competitive packing, challenging the assumption that species richness always increases ecosystem function [2]. |
| Trait-Based: Co-Selection [25] [26] | Exposure to sub-inhibitory concentrations of heavy metals (e.g., Cu, Zn) or biocides (e.g., QACs) selected for bacterial populations with increased antibiotic resistance. | Non-antibiotic environmental contaminants can co-select for antibiotic resistance via co-resistance (linked genes) or cross-resistance (e.g., efflux pumps), highlighting a major risk factor in AMR proliferation [25] [26]. |
| Directed Evolution: General [3] | A single round of error-prone PCR typically creates libraries of 10^6 - 10^10 variants. FACS-based screening can sort >10^8 cells per hour. | Enables exploration of a vast sequence space that is impossible with rational design, allowing for the discovery of unexpected solutions. |
| Directed Evolution: RACHITT [3] | Method yielded a 15-fold increase in crossover frequency compared to earlier DNA shuffling techniques. | Results in higher diversity libraries and more efficient exploration of chimeric protein sequences. |
The Scientist's Toolkit: Research Reagent Solutions
Essential materials and reagents for implementing these evolutionary approaches.
| Item | Function & Application |
|---|---|
| Error-Prone PCR Kit | Utilizes a DNA polymerase with low fidelity and biased reaction conditions to introduce random point mutations during gene amplification for directed evolution library generation [3]. |
| Phage Display Library | A collection of filamentous phages, each displaying a different protein variant on its surface. Used for high-throughput selection of high-affinity binders from libraries exceeding 10^9 members [3]. |
| FACS (Fluorescence-Activated Cell Sorter) | An instrument that sorts individual cells based on fluorescence. In directed evolution, it is used to isolate enzyme or binder variants labeled with a fluorescent product or tag [3]. |
| Functional Trait Database | Curated databases (e.g., TRY for plant traits) that provide standardized trait measurements. Used in trait-based studies for large-scale comparative analyses and modeling [24]. |
| Metals & Biocides | Heavy metals (e.g., Copper, Zinc) and biocides (e.g., Quaternary Ammonium Compounds) are used in co-selection studies to apply selective pressure and investigate the evolution of cross-resistance in microbial communities [25] [26]. |
| PROTAC SOS1 degrader-5 | PROTAC SOS1 degrader-5, MF:C45H51F3N8O7, MW:872.9 g/mol |
| (1S,9R)-Exatecan mesylate | (1S,9R)-Exatecan mesylate, MF:C25H26FN3O7S, MW:531.6 g/mol |
Workflow and Conceptual Diagrams
The diagrams below illustrate the logical flow of each evolutionary approach.
Trait-Based Eco-Evolutionary Framework
Directed Evolution Workflow
The relationship between a protein's amino acid sequence and its biological function is one of the most fundamental yet poorly understood aspects of molecular biology. Despite decades of research, accurately predicting function from sequence alone remains exceptionally difficult, creating a significant bottleneck in fields ranging from drug development to enzyme engineering. This challenge stems from the highly complex nature of sequence-function relationships, which arises from the intricate interplay between biophysical constraints and evolutionary history [27]. The astronomical vastness of sequence space further complicates this picture—a typical protein several hundred amino acids long represents 20³â°â° possible sequences, a number that exceeds the total atoms in the universe [28]. This review examines why this prediction problem persists, comparing two dominant approaches for navigating this complexity: trait-based rational design and empirical directed evolution. We objectively evaluate their performance through experimental data and methodological analysis, providing researchers with a framework for selecting appropriate strategies for protein engineering challenges.
The Core Mechanisms Underlying Prediction Challenges
Epistasis and Context-Dependent Effects
A primary source of prediction difficulty lies in epistasis, where the effect of a mutation depends on its genetic context. Rather than acting independently, amino acid residues engage in complex, higher-order interactions that profoundly influence protein function [28]. Traditional reference-based analyses, which measure mutational effects relative to a single wild-type sequence, often overestimate epistasis by propagating measurement noise and local idiosyncrasies into high-order interactions [29]. This can lead to unnecessarily complex descriptions of genetic architecture. Reference-free analysis (RFA) addresses this by taking a global perspective, defining amino acid effects relative to the average across all possible sequences rather than a single reference point [29]. Studies implementing RFA reveal that sequence-function relationships are surprisingly simple in many cases, with context-independent amino acid effects and pairwise interactions explaining over 92% of phenotypic variance across 20 experimental datasets [29].
Rugged Fitness Landscapes and Nonlinearity
The presence of epistasis creates rugged fitness landscapes rather than smooth, easily navigable surfaces. These landscapes are characterized by multiple peaks, valleys, and plateaus, making it difficult to predict the functional outcome of multiple simultaneous mutations [28]. This ruggedness arises not only from direct structural contacts between residues but also from indirect effects modulated by ligands, substrates, allostery, cofactors, and conformational dynamics [28]. Additionally, global nonlinearities in the relationship between sequence and function further complicate prediction. Without accounting for these nonlinear transformations, analyses must invoke pervasive complex interactions to explain why mutation effects vary across genetic backgrounds [29]. For example, a stability threshold effect can occur where individually beneficial but destabilizing mutations combine to completely abolish activity [28].
Sparse Determinants and Multi-Functionality
Protein sequence-function relationships exhibit sparse determinism, where a minute fraction of possible amino acids and interactions account for the majority of functional variance [29]. This sparsity makes identification of key determinants challenging, especially with limited experimental data. Furthermore, many proteins exhibit multi-functionality, performing multiple distinct biological roles. The MoonProt database catalogs hundreds of proteins experimentally demonstrated to have more than one function [30]. This multi-functionality often arises from small sequence or structural changes that are difficult to predict a priori. For instance, the enolase superfamily contains evolutionarily related enzymes with similar TIM-barrel folds that catalyze different reactions, including enolases, muconate lactonizing enzymes, and mandelate racemases [30]. Small changes in active site residues are sufficient to alter catalytic specificity, creating prediction challenges even among well-characterized protein families.
Comparative Analysis of Engineering Approaches
Trait-Based Rational Design
Trait-based approaches rely on prior structural and mechanistic knowledge to make informed predictions about which sequence changes will produce desired functions. These methods leverage detailed understanding of protein biochemistry to design variants with improved or altered properties.
Methodological Framework
Trait-based design typically begins with structural analysis to identify key residues influencing target functions, such as active site residues for catalysis or interface residues for binding interactions. Researchers then employ computational modeling to predict the functional consequences of mutations, often using molecular dynamics simulations or energy calculations. Finally, a limited set of designed variants is synthesized and experimentally validated [30]. This approach requires high-quality structural data and robust computational models that accurately capture sequence-structure-function relationships.
Experimental Performance and Limitations
While successful in many applications, trait-based design faces significant limitations. Performance varies considerably depending on the protein system and target function, with particularly poor outcomes for functions lacking clearly correlated structural features [30]. For example, predicting protein-protein interaction sites remains challenging because these interfaces often consist of relatively smooth surface regions without well-conserved motifs [30]. Similarly, many RNA-binding proteins lack canonical RNA-binding domains, making computational prediction of interaction sites difficult. These limitations underscore the fundamental gaps in our understanding of how sequence encodes function.
Directed Evolution Approaches
Directed evolution mimics natural selection in the laboratory, using iterative rounds of diversification and selection to improve protein functions without requiring detailed mechanistic knowledge.
Methodological Framework
The directed evolution cycle begins with the creation of genetic diversity through random mutagenesis, DNA shuffling, or targeted methods like CRISPR-Cas systems [31] [32]. This variant library is then subjected to high-throughput screening or selection to identify individuals with improved functional characteristics. The top-performing variants serve as templates for subsequent rounds of diversification and selection, progressively optimizing the desired function [28] [32]. Unlike trait-based approaches, directed evolution does not require prior structural knowledge, instead relying on functional screening to guide the optimization process.
Experimental Performance and Applications
Directed evolution has demonstrated remarkable success across diverse applications. In agricultural biotechnology, researchers used CRISPR-Cas-mediated directed evolution to develop herbicide-resistant crops by evolving rice acetyl-coenzyme A carboxylase (ACC) and acetolactate synthase (ALS1) variants [31]. These evolved enzymes conferred resistance while maintaining catalytic function, with specific mutations (P1927F and W2125C in ACC; P171F in ALS1) identified as responsible for the herbicide-tolerant phenotype [31]. Similarly, directed evolution of rice splicing factor SF3B1 generated variants (SGR mutants) resistant to splicing inhibitors herboxidiene and pladienolide B while maintaining splicing functionality [31]. These successes highlight the power of directed evolution to optimize complex functions without detailed structural knowledge.
Machine Learning-Enabled Approaches
Recent advances integrate machine learning with directed evolution, creating hybrid approaches that leverage large-scale sequence-function data.
Methodological Framework
Machine learning methods for protein engineering typically involve training models on deep mutational scanning (DMS) data, which combines high-throughput functional screening with next-generation sequencing to map sequence-function relationships for thousands to millions of variants [33]. These models learn the mapping between sequence and function, enabling prediction of functional outcomes for uncharacterized sequences. Positive-unlabeled learning frameworks have been developed specifically to handle DMS data, which often lacks explicit negative examples [33]. The trained models guide subsequent library design, creating an iterative optimization cycle that reduces experimental burden.
Experimental Performance and Applications
Machine learning approaches have demonstrated excellent predictive performance across diverse protein systems. In one notable application, researchers used learned sequence-function models to design highly stabilized enzymes, successfully extrapolating beyond the training data to create thermostable variants [33]. These methods effectively identify key functional determinants while handling the high dimensionality and correlations inherent to protein sequence data. The GOLabeler method, which integrates multiple data sources using machine learning, significantly outperformed other function prediction algorithms in the critical assessment of functional annotation (CAFA) challenges [30].
Table 1: Comparative Performance of Protein Engineering Approaches
| Approach | Key Methodologies | Typical Experimental Throughput | Success Rate | Primary Limitations |
|---|---|---|---|---|
| Trait-Based Rational Design | Structure-based design, computational modeling, molecular dynamics | Low (10-100 variants) | Variable (high for well-characterized systems) | Requires detailed structural knowledge; poor for complex functions |
| Directed Evolution | Random mutagenesis, DNA shuffling, CRISPR-Cas diversification | Medium to High (10³-10⸠variants) | Consistently high across diverse systems | Limited by screening capacity; can require many iterations |
| Machine Learning-Enabled | Deep mutational scanning, positive-unlabeled learning, neural networks | Very High (10âµ-10â¹ variants in silico) | Increasingly high with sufficient training data | Requires large training datasets; model interpretability challenges |
Table 2: Quantitative Performance Metrics Across Representative Studies
| Protein System | Engineering Goal | Trait-Based Results | Directed Evolution Results | Machine Learning Results |
|---|---|---|---|---|
| Rice SF3B1 [31] | Herbicide resistance | N/A | SGR mutants with full splicing function and drug resistance | N/A |
| Acetyl-coenzyme A carboxylase [31] | Herbicide resistance | N/A | P1927F and W2125C mutations conferring haloxyfop resistance | N/A |
| GB1 binding protein [32] | Enhanced binding affinity | N/A | N/A | Accurate prediction of optimized combinatorial libraries |
| Multiple enzyme systems [33] | Thermostability | Variable success | N/A | Successful design of stabilized enzymes |
Experimental Protocols for Key Methodologies
Deep Mutational Scanning Protocol
Deep mutational scanning (DMS) provides comprehensive sequence-function maps by combining high-throughput functional screening with deep sequencing [33]. The protocol begins with library generation, creating variant libraries covering targeted regions through saturation mutagenesis, error-prone PCR, or oligonucleotide synthesis. The library is then subjected to functional screening, applying selective pressure to separate functional from non-functional variants. This is followed by high-throughput sequencing of pre-selection and post-selection populations to quantify variant enrichment. Finally, enrichment analysis calculates fitness scores for each variant based on frequency changes, generating a quantitative sequence-function map [33]. DMS data typically contains only positive and unlabeled examples, necessitating specialized computational approaches like positive-unlabeled learning for accurate modeling [33].
CRISPR-Cas-Mediated Directed Evolution Protocol
CRISPR-Cas systems enable targeted diversification for directed evolution in native genomic contexts [31]. The protocol involves sgRNA library design to target specific gene regions, library delivery into plant cells via Agrobacterium-mediated transformation, and CRISPR-Cas editing to generate diverse mutations through non-homologous end joining (NHEJ) or homology-directed repair (HDR). Edited cells are then subjected to selection pressure (e.g., herbicide application), followed by recovery and analysis of resistant variants [31]. This approach was successfully used to evolve herbicide resistance in rice by targeting the SF3B1 gene, generating splicing factor variants that maintained function while gaining resistance to pladienolide B and herboxidiene [31].
Reference-Free Analysis Protocol
Reference-free analysis (RFA) addresses limitations of traditional reference-based approaches by providing a global perspective on sequence-function relationships [29]. The method involves data collection from combinatorial mutagenesis studies, global mean calculation across all variants, first-order effect estimation for each amino acid state as the difference between sequences containing that state and the global mean, and epistatic effect calculation for state combinations as differences between observed and expected means based on lower-order effects [29]. RFA can be accurately estimated using least-squares regression even with missing data and provides the most efficient explanation of sequence-function relationships by maximizing variance explained at each epistatic order [29].
Visualizing Sequence-Function Relationship Concepts
Protein Fitness Landscape Complexity
Directed Evolution Workflow
Reference-Based vs Reference-Free Analysis
The Scientist's Toolkit: Essential Research Reagents and Solutions
Table 3: Key Research Reagents for Sequence-Function Studies
| Tool/Category | Specific Examples | Function/Application | Key Considerations |
|---|---|---|---|
| Diversification Technologies | Error-prone PCR, DNA shuffling, MAGE, CRISPR-Cas systems | Generate genetic variation for functional screening | Mutation rate, library diversity, bias introduction |
| Screening Systems | FACS, microfluidics, growth selection, phage display | Identify functional variants from libraries | Throughput, sensitivity, false positive/negative rates |
| Sequence-Function Mapping | Deep mutational scanning, phage-assisted continuous evolution | Comprehensive analysis of variant effects | Coverage, quantitative accuracy, statistical power |
| Computational Tools | Positive-unlabeled learning, reference-free analysis, neural networks | Model sequence-function relationships | Data requirements, interpretability, predictive accuracy |
| Specialized Databases | UniProt, PDB, MoonProt, DisProt, Enzyme Portal | Access to sequence, structure, and function data | Coverage, annotation quality, update frequency |
| 2'-Deoxyadenosine-13C10 | 2'-Deoxyadenosine-13C10, MF:C10H13N5O3, MW:261.17 g/mol | Chemical Reagent | Bench Chemicals |
| Me-Tet-PEG5-COOH | Me-Tet-PEG5-COOH, MF:C24H35N5O8, MW:521.6 g/mol | Chemical Reagent | Bench Chemicals |
The relationship between protein sequence and function remains difficult to predict a priori due to epistasis, rugged fitness landscapes, and sparse functional determinants. Our comparative analysis reveals that while trait-based rational design provides mechanistic insights for well-characterized systems, directed evolution offers more consistent success across diverse protein engineering challenges. Machine learning approaches are rapidly bridging this divide, leveraging large-scale experimental data to build predictive models that capture complex sequence-function relationships. The emerging paradigm integrates these approaches, using directed evolution to generate functional data and machine learning to extract generalizable principles. This integrated framework promises to accelerate protein engineering for therapeutic development, industrial applications, and fundamental biological research, gradually transforming the sequence-function relationship from a fundamental mystery to a tractable engineering problem.
Toolkits and Techniques: Implementing Directed Evolution and Trait Analysis
Genetic diversification is a cornerstone of biological research and biotechnology, enabling the exploration of gene function and the evolution of proteins with novel traits. The methods to achieve this diversification broadly fall into two categories: random, untargeted approaches (like error-prone PCR and DNA shuffling) and precise, targeted approaches (exemplified by CRISPR-Cas systems). This guide provides an objective comparison of these three key techniques—Error-Prone PCR, DNA Shuffling, and CRISPR-Cas Systems—framed within the broader thesis of trait-based versus directed evolution approaches for optimizing biological functions. We summarize their mechanisms, applications, and experimental data to inform researchers and drug development professionals in selecting the appropriate tool for their community optimization research.
The following table offers a high-level comparison of the three genetic diversification methods.
| Feature | Error-Prone PCR | DNA Shuffling | CRISPR-Cas Systems |
|---|---|---|---|
| Core Principle | Introduces random point mutations during PCR amplification using error-prone conditions [34]. | Recombines fragments from related DNA sequences in vitro to create chimeric genes [35]. | Uses a programmable RNA-guided Cas nuclease to make targeted double-strand breaks in the genome, engaging cellular repair mechanisms to introduce changes [36] [37]. |
| Primary Type of Diversity | Point mutations, small insertions/deletions [34]. | Recombination of existing mutations and gene homologs; can create novel combinations of sequences from parent genes [35]. | User-defined mutations via HDR; random indels via NHEJ; can be targeted to specific chromosomal loci [36]. |
| Typical Library Size | Limited by cloning efficiency [34]. | Can rapidly generate very large libraries (e.g., millions of clones) [35]. | Limited by HDR efficiency in many systems, but sgRNA libraries can offer extensive coverage [36]. |
| Key Advantage | Technically simple, no structural knowledge of protein required. | Rapidly recombines beneficial mutations from multiple parents. | Enables precise, chromosomal diversification at the native locus, preserving endogenous regulation [36]. |
| Major Limitation | Mutations are random and largely blind to function. | Requires sequence homology or common restriction sites for some methods [35]. | Lower efficiency of HDR in mammalian cells; risk of unintended on-target structural variations [36] [38]. |
| Tyk2-IN-15 | Tyk2-IN-15|Potent TYK2 Inhibitor|For Research | Bench Chemicals | |
| Ebov-IN-6 | EBOV-IN-6 | EBOV-IN-6 is a benzothiazepine compound with anti-Ebola virus (EBOV) research activity (IC50 = 10 μM). This product is for research use only and not for human use. | Bench Chemicals |
Experimental Protocols and Workflows
Below are the standardized experimental workflows for each method, from library generation to variant screening.
Error-Prone PCR Workflow
- Step 1: Library Generation. The gene of interest is amplified by PCR under "sloppy" conditions. This involves using a polymerase with low fidelity, increasing the concentration of MgClâ‚‚, adding MnClâ‚‚, and using unequal concentrations of the four dNTPs. These conditions promote misincorporation of nucleotides during amplification, resulting in a pool of PCR products with random point mutations [34].
- Step 2: Cloning and Transformation. The mutated PCR products are then cloned into a suitable plasmid expression vector. This library of plasmids is transformed into a bacterial host (e.g., E. coli) to create a library of clones, each expressing a different variant of the gene [34].
- Step 3: Screening/Selection. The transformed library is screened using high-throughput assays (e.g., for enzyme activity, ligand binding, or fluorescence) to identify clones with the desired improved or altered function [34].
DNA Shuffling Workflow
- Step 1: Fragmentation. Multiple parent genes (which can be natural homologs or pre-mutated variants) are physically fragmented. This can be done enzymatically using DNase I [35] [39] or via more controlled methods like Nucleotide Exchange and Excision Technology (NExT), which uses the incorporation of uracil and subsequent excision to generate fragments [39].
- Step 2: Reassembly. The fragments are mixed and subjected to a primer-less PCR. Due to sequence homologies, fragments from different parents can anneal to each other. The DNA polymerase then extends these overlapping fragments, reassembling them into full-length, chimeric genes [35].
- Step 3: Cloning and Selection. The reassembled genes are amplified with primers and cloned into an expression vector. The resulting library, which contains hybrids recombining sequences from all parent genes, is then screened for variants with enhanced or novel properties [35].
CRISPR-Cas Mediated Chromosomal Diversification Workflow
- Step 1: Component Design. A single-guide RNA (sgRNA) is designed to target the specific genomic locus of interest. A library of donor DNA templates (HDR templates) is synthesized, containing the desired diversified sequences (e.g., saturated mutations, barcodes, or coding variants) flanked by homologous arms [36].
- Step 2: Delivery and Cleavage. The sgRNA, Cas9 nuclease, and the HDR template library are co-delivered into the target cells. The Cas9-sgRNA complex creates a precise double-strand break (DSB) in the chromosome at the target site [36] [37].
- Step 3: Homology-Directed Repair. The cell's repair machinery uses the supplied donor DNA library as a template for HDR, thereby integrating the diversified sequences directly into the native chromosomal location. This ensures the variants are expressed under endogenous regulatory control [36].
- Step 4: Functional Screening. The population of edited cells, now a library of genetic variants at the target locus, can be subjected to a selective pressure (e.g., a drug, growth condition, or fluorescent assay). Variants with enhanced function are enriched, and their sequences can be identified via next-generation sequencing [36].
Performance and Quantitative Data Comparison
The tables below summarize key performance metrics and experimental data for these methods, illustrating their practical capabilities and limitations.
Table 1: Key Performance Metrics for Genetic Diversification Methods
| Method | Diversity Introduced | Typical Mutation Rate/ Efficiency | Key Applications |
|---|---|---|---|
| Error-Prone PCR | Point mutations (transitions, transversions), small indels [34]. | Error rate can be tuned up to ~2% per base [34]. | Evolving individual proteins for improved stability, activity, or altered substrate specificity [34]. |
| DNA Shuffling | Recombination of point mutations and homologous blocks from multiple parent genes [35]. | Can generate libraries of >10ⴠto 10ⶠclones; low background mutation rate (e.g., 0.1% with NExT) [39]. | Rapidly combining beneficial mutations from different homologs; family shuffling to evolve complex traits like herbicide detoxification [35]. |
| CRISPR-Cas Systems | User-defined single nucleotide variants (SNVs) or small sequences integrated via HDR; random indels via NHEJ [36]. | HDR efficiency is variable and often low (e.g., 0.2% - 3.3% in mammalian cells); can be increased with small molecules, but this may raise SV risk [36] [38]. | Saturation genome editing to map variant effects; CasPER for directed evolution of pathways (11-fold yield increase reported); gene therapy [36] [38]. |
Table 2: Documented Experimental Outcomes from Literature
| Method | Experimental System | Reported Outcome | Reference & Context |
|---|---|---|---|
| DNA Shuffling | Chloramphenicol acetyltransferase (CAT) gene | Successful shuffling of truncated CAT variants with an average parental fragment size of 86 bp and a low mutation rate (0.1%). | NExT DNA shuffling methodology [39]. |
| CRISPR-HDR (Saturation Editing) | Human HAP1 cells | Efficiency of saturation editing for exons in genes like BRCA1 ranged from 0.2% to 3.33%. | Functional profiling of disease-associated genes [36]. |
| CRISPR-HDR (CasPER) | Yeast Mevalonate Pathway | Integration of 300-600 bp donor sequences with >98% efficiency, leading to an 11-fold increase in isoprenoid production after selection. | Directed evolution of endogenous yeast genes [36]. |
Critical Considerations for Research Applications
Safety and Limitations: The Case of CRISPR-Cas
While revolutionary, CRISPR-Cas systems carry specific risks that must be accounted for in experimental design, especially for therapeutic applications. Beyond well-known off-target effects, a pressing concern is the introduction of on-target structural variations (SVs). These include large deletions (kilobase- to megabase-scale), chromosomal translocations, and other complex rearrangements [38]. Critically, strategies to improve HDR efficiency, such as using DNA-PKcs inhibitors (e.g., AZD7648), can dramatically increase the frequency of these SVs—by a thousand-fold for some translocations [38]. Furthermore, traditional short-read amplicon sequencing often fails to detect these large deletions, leading to an overestimation of precise HDR efficiency [38]. Therefore, comprehensive genomic integrity checks using long-read sequencing or dedicated SV-detection assays (like CAST-Seq) are recommended for rigorous safety assessment [38].
Integration in Trait-Based vs. Directed Evolution Frameworks
The choice of diversification method aligns with different ecological optimization strategies:
- Trait-Based Approaches leverage rational design by assembling communities from members with known, complementary traits. This is analogous to using CRISPR for precise, hypothesis-driven saturation editing to understand gene function [36] [9].
- Directed Evolution Approaches remain agnostic to mechanism, instead using iterative rounds of diversification and selection to find high-performing variants or communities. Error-prone PCR and DNA shuffling are classic tools for this, mimicking the evolutionary process of mutation and recombination [9] [34] [35]. CRISPR methods like CasPER now bring the power of targeted, chromosomal diversification to this paradigm [36].
Essential Research Reagent Solutions
The following table lists key reagents and their functions crucial for implementing these genetic diversification methods.
Table 3: Key Reagents for Genetic Diversification Experiments
| Reagent / Tool | Function | Example Use Cases |
|---|---|---|
| Taq Polymerase & Sloppy Buffer | Enzyme and optimized buffer system for performing error-prone PCR. Contains elevated Mg²âº, Mn²âº, and unbalanced dNTPs to promote nucleotide misincorporation [34]. | Random mutagenesis of a gene to create a variant library for enzyme evolution. |
| DNase I or NExT Reagents | Enzymes for fragmenting DNA. DNase I randomly cleaves DNA, while NExT (using dUTP incorporation, UDG, and piperidine) offers a more controllable fragmentation pattern [35] [39]. | Initial step in DNA shuffling to create fragments for recombination. |
| Cas9 Nuclease (Wild-type & HiFi) | Effector protein that creates a double-strand break at a DNA target specified by the sgRNA. High-fidelity (HiFi) variants reduce off-target editing [36] [38] [37]. | Targeted chromosomal gene disruption (KO) or diversification via HDR. |
| sgRNA Library | A pooled collection of single-guide RNAs designed to target multiple genomic sites or to cover a specific gene region comprehensively [36]. | Genome-wide screens or saturation editing of a specific gene. |
| HDR Donor Template Library | A pool of single or double-stranded DNA oligonucleotides containing the variant sequences to be integrated, flanked by homology arms matching the target locus [36]. | Introducing a library of specific mutations (e.g., all possible amino acid changes) into a genomic locus. |
| DNA-PKcs Inhibitor (e.g., AZD7648) | Small molecule inhibitor of the NHEJ DNA repair pathway. Used to tilt the balance of DSB repair towards HDR, thereby increasing editing efficiency [38]. | Use with caution. Boosts HDR rates but significantly increases risk of large structural variations [38]. |
Error-Prone PCR, DNA Shuffling, and CRISPR-Cas Systems each offer distinct pathways for genetic diversification, catering to different research needs. The choice between them hinges on the core objective: whether the aim is broad, random exploration of sequence space or precise, targeted hypothesis testing. As the field of synthetic biology and community optimization advances, the integration of robust traditional methods like DNA shuffling with the precision of CRISPR-Cas systems promises to be a powerful strategy. This hybrid approach can leverage the strengths of both directed evolution and rational, trait-based design to engineer biological systems with unprecedented control and functionality. Researchers must carefully weigh the trade-offs in precision, efficiency, and safety—particularly the risk of structural variations with CRISPR—when designing their experimental pipelines.
For decades, random mutagenesis served as the cornerstone of protein engineering, mimicking natural evolution through untargeted genetic diversification and phenotypic screening. While successful, this approach explores sequence space inefficiently, often requiring immense screening efforts and frequently failing to identify optimal mutations due to its undirected nature. The field has progressively shifted towards more precise, knowledge-driven strategies. Site-saturation mutagenesis (SSM) and other targeted evolution methodologies now empower researchers to conduct systematic, focused investigations into protein function and stability, leading to more efficient engineering of biocatalysts, therapeutics, and research tools [40] [41]. This guide objectively compares the performance of these targeted strategies against classical random mutagenesis, providing a framework for selecting the optimal approach for community optimization and drug development research.
Methodological Comparison: Core Techniques and Workflows
Fundamental Principles
- Random Mutagenesis introduces mutations randomly throughout a gene or genome using methods like error-prone PCR or chemical mutagens. It requires no prior structural or mechanistic knowledge but demands high-throughput screening to find rare beneficial mutations amid predominantly neutral or deleterious ones [42].
- Site-Saturation Mutagenesis (SSM) is a targeted technique that systematically substitutes a single codon (or set of codons) with all possible amino acids at that position. This creates a comprehensive library for a specific region, allowing researchers to exhaustively map the sequence-function relationship at defined sites [43] [41].
- Semi-Rational Design leverages bioinformatics and structural biology data to identify key "hotspot" residues for mutagenesis. This approach uses information from multiple sequence alignments, phylogenetic analysis, or protein structures to design small, functionally rich libraries, thereby minimizing screening effort while maximizing the potential for improvement [40].
Experimental Workflows
The fundamental difference between these strategies is visualized in their experimental pathways. The diagram below illustrates the key steps involved in random mutagenesis versus the more streamlined site-saturation and semi-rational approaches.
Performance and Outcome Comparison
The theoretical advantages of targeted strategies are borne out in direct experimental comparisons. The following table summarizes key performance metrics and documented outcomes.
Table 1: Comparative Analysis of Mutagenesis Strategies
| Feature | Random Mutagenesis | Site-Saturation Mutagenesis (SSM) | Semi-Rational Design |
|---|---|---|---|
| Library Size | Very large (often >10^6 variants) [40] | Defined by design (e.g., 19-500 variants per site) [40] [44] | Small, focused (often <1000 variants) [40] |
| Prior Knowledge Required | None | Protein sequence | Protein sequence, structure, and/or mechanism |
| Control Over Mutations | None | Precise control over targeted positions | Precise control over targeted positions and diversity |
| Typical Screening Effort | Very high | Low to moderate | Low |
| Functional Content | Low (many neutral/deleterious mutations) | High at targeted site | Very high |
| Best Use Case | No structural data; exploring vast sequence space | Identifying key residues; engineering active sites | Stability, specificity, and multi-property optimization |
| Reported Outcome Example | General strain improvement [42] | 200-fold improved activity & 20-fold enantioselectivity in an esterase [40] | 32-fold improved activity in a dehalogenase; redesigned transaminase for industrial process [40] |
Large-scale studies demonstrate the power of SSM. A 2025 study performing SSM on over 500 human protein domains quantified the effects of more than 500,000 missense variants, revealing that a majority of pathogenic variants reduce protein stability and providing an immense dataset for clinical variant interpretation and computational tool benchmarking [45].
Experimental Protocols for Key Techniques
One-Step Site-Saturation Mutagenesis Protocol
This is a widely used PCR-based method for introducing mutations at a single residue [46] [43].
- Primer Design: Design two complementary primers that are partially overlapping and contain the degenerate codon (e.g., NNK or NNS, where K = G/T and S = G/C) at the target site. These primers anneal to the same sequence on opposite strands.
- PCR Amplification: Perform a PCR using a high-fidelity DNA polymerase with the plasmid template and the mutagenic primers. This method results in linear amplification.
- Template Digestion: Digest the PCR product with DpnI restriction enzyme, which specifically cleaves methylated DNA, to eliminate the original parental plasmid template.
- Transformation: Transform the digested PCR product into competent E. coli cells. The cellular machinery repairs the nicks in the plasmid, resulting in a library of mutant plasmids.
Two-Step Megaprimer PCR Protocol
For "difficult-to-randomize" genes (e.g., long, high GC-content), a two-step megaprimer method proves superior, generating higher-quality libraries [46].
- First PCR - Megaprimer Generation: Perform a PCR using one mutagenic primer (with an NNK codon) and one non-mutagenic (silent) primer. Use the target plasmid as a template to generate a short, mutated DNA fragment.
- Product Purification: Purify the resulting DNA fragment, which serves as a megaprimer.
- Second PCR - Whole-Plasmid Amplification: Use the purified megaprimer in a second PCR to amplify the entire plasmid.
- Template Digestion and Transformation: Digest the final PCR product with DpnI and transform into E. coli, as in the one-step protocol.
The Scientist's Toolkit: Essential Research Reagents
Successful implementation of these strategies relies on a suite of specialized reagents and computational tools.
Table 2: Key Reagents and Tools for Targeted Evolution
| Category | Item | Function | Example Use Case |
|---|---|---|---|
| Molecular Biology | NNK/NNS Degenerate Codons | Encodes all 20 amino acids with a single stop codon [43]. | General-purpose site-saturation mutagenesis. |
| Non-Degenerate Codon Mixes | Defines a specific, reduced amino acid set; avoids stop codons [41]. | Creating "smart" libraries with reduced screening burden. | |
| High-Fidelity DNA Polymerase | Accurate PCR amplification with low error rates. | All PCR-based mutagenesis protocols. | |
| DpnI Restriction Enzyme | Selective digestion of methylated parental plasmid template. | Enriching for newly synthesized mutant DNA after PCR. | |
| Computational Tools | 3DM Database | Analyzes protein superfamilies to identify evolutionarily allowed substitutions [40]. | Guiding semi-rational library design for enantioselectivity. |
| HotSpot Wizard | Identifies mutable positions based on sequence and structure data [40]. | Engineering catalytic activity and stability. | |
| RosettaDesign Software | Models and designs protein structures and sequences. | De novo enzyme design and active site remodeling [40]. | |
| Screening & Selection | Fluorescence-Activated Cell Sorting (FACS) | High-throughput isolation of cells based on fluorescent markers. | Screening large libraries for binding or enzymatic activity. |
| Protein Fragment Complementation Assay (aPCA) | Links protein abundance/solubility to cell growth or survival [45]. | Large-scale stability profiling of missense variants. | |
| Igermetostat | Igermetostat|EZH2 Inhibitor|CAS 2409538-60-7 | Igermetostat is a potent EZH2 inhibitor for cancer research (in vivo/vitro). For Research Use Only. Not for human or veterinary use. | Bench Chemicals |
| Paynantheine-d3 | Paynantheine-d3, MF:C23H28N2O4, MW:399.5 g/mol | Chemical Reagent | Bench Chemicals |
Strategic Application in Research and Development
The choice of mutagenesis strategy should be dictated by the specific research goal and the available information about the protein system. The decision logic for selecting the most appropriate method is summarized below.
For drug development, SSM is invaluable for humanizing antibodies, optimizing biologic stability, and understanding drug resistance mechanisms by comprehensively probing binding interfaces. In enzyme engineering for industrial processes, semi-rational design has successfully created biocatalysts with enhanced stereoselectivity, thermostability, and organic solvent tolerance [40]. Furthermore, SSM has been used to challenge established enzyme mechanisms, leading to revised models of catalysis that can inform the design of more effective inhibitors [44].
Emerging strategies are also moving beyond simple top-variant selection. Computational models suggest that splitting populations and using tuned "selection functions" during directed evolution can improve outcomes by maintaining diversity and helping populations escape local fitness maxima, potentially increasing the probability of finding the global optimum by up to 19-fold [20] [47].
High-throughput screening (HTS) technologies are indispensable in modern biological research and drug development, enabling the rapid evaluation of vast molecular or cellular libraries. Within the broader thesis of community optimization research, these tools offer distinct pathways: trait-based approaches, which rely on screening for predefined, measurable characteristics, and directed evolution approaches, which use iterative selection and diversification to evolve desired functions. This guide objectively compares the performance of three cornerstone HTS technologies—Flow Cytometry and Cell Sorting (FACS), Display Technologies, and Biosensors—in the context of these two research philosophies.
The following table summarizes the core characteristics, performance metrics, and ideal applications of each technology.
| Feature | FACS (Fluorescence-Activated Cell Sorting) | Display Technologies (e.g., Phage Display) | Biosensors (Genetically Encoded) |
|---|---|---|---|
| Core Principle | Physical separation of cells/particles based on optical properties (light scattering & fluorescence) [48] [49]. | Physical coupling between a displayed protein (e.g., antibody, peptide) and its genetic code, enabling library screening [50]. | Intracellular conversion of target metabolite concentration into a quantifiable optical signal (e.g., fluorescence) [51]. |
| Primary Application | Analysis and isolation of cell subpopulations; multiparameter single-cell analysis. | Identification of high-affinity ligands (peptides, antibodies) against specific targets (e.g., cancer biomarkers) [50]. | Real-time, in situ monitoring of metabolite production and high-throughput screening of microbial cell factories [51]. |
| Throughput | High (up to 70,000 events/second for analysis) [52]. | Very High (can screen libraries containing billions of clones). | Extremely High (allows in situ screening of colonies on agar plates) [51]. |
| Quantitative Data Output | Multiplexed fluorescence intensity, cell size, granularity. | Binding affinity (e.g., KD), selectivity. | Fluorescence intensity correlated to metabolite concentration. |
| Sensitivity | High-sensitivity detection of dim fluorescent populations [48] [49]. | Capable of identifying rare, high-affinity binders from large libraries. | Sensitive enough to detect endogenous production of metabolites like 5-aminolevulinic acid [51]. |
| Key Instrument/System | BD FACSAria Fusion, BD FACSAria III [48] [49]. | Phage display libraries (peptide, scFv, nanobody). | Whole-cell biosensors with engineered transcription factors. |
| Thesis Context: Trait-Based Screening | Excellent for sorting based on measurable traits like surface marker expression (e.g., isolating Treg cells based on FoxP3) [49]. | Limited direct application; primarily for probe discovery. | Not applicable. |
| Thesis Context: Directed Evolution | Used downstream to isolate variants with improved traits (e.g., fluorescence, binding) from diversified libraries. | The core technology for iterative affinity maturation of proteins [50]. | Enables direct high-throughput screening of engineered metabolic pathways for optimized production [51]. |
Experimental Protocols and Workflows
Protocol: Cell Sorting using a BD FACS System (e.g., BD FACSAria Fusion)
This protocol details the process for high-speed, high-purity sorting of specific cell populations, a key step in both trait-based selection and directed evolution workflows [48] [52].
- Sample Preparation: Cells are stained with fluorescently conjugated antibodies or dyes targeting specific markers (e.g., surface receptors). The sample is resuspended in a suitable buffer, filtered to remove clumps, and kept on ice.
- Instrument Setup & Quality Control: Lasers and fluidics are initialized. Daily quality control is performed using calibration beads to ensure optimal laser alignment and fluorescence detection sensitivity. The instrument's fixed alignment cuvette flow cell minimizes startup time and variability [48] [49].
- Drop-Delay Calculation: Using BD FACS Accudrop Technology, the drop-delay (the time between droplet formation and charging) is automatically calculated and maintained, which is critical for sort accuracy [48] [49].
- Acquisition & Gating: A small portion of the sample is run to establish populations of interest. Sequential "gates" are set on scatter and fluorescence plots to define the target cell population with high purity.
- Sorting: The instrument is set to sort mode. Defined cells are charged and deflected into collection tubes (e.g., 15 mL) or multi-well plates (e.g., 96-well). The collection chamber can be temperature-controlled (cooling or heating) [52].
- Post-Sort Analysis: A sample of the sorted cells is re-analyzed to confirm purity, which is typically >95% [49].
High-Level FACS Sorting Workflow
Protocol: Developing a Whole-Cell Biosensor for Metabolite Detection
This protocol outlines the creation of a genetically encoded biosensor for high-throughput screening, a prime tool for directed evolution of metabolic pathways [51].
- Transcription Factor (TF) Selection & Engineering: A native TF (e.g., AsnC, which responds to L-asparagine) is selected as a scaffold based on structural similarity to the target metabolite (5-aminolevulinic acid or 5-ALA). Saturation mutagenesis is performed on key amino acid residues in the TF's ligand-binding domain to alter its specificity.
- Library Construction & Screening: The mutant TF library is cloned into a plasmid to control the expression of a reporter gene, such as the red fluorescent protein (RFP). The library is transformed into E. coli.
- Positive-Negative Screening:
- Positive Selection: Colonies are grown on agar plates containing the target metabolite (5-ALA). Mutants that successfully respond to 5-ALA and produce RFP (visible as red colonies) are selected.
- Negative Selection: These potential hits are then replica-plated onto media containing the original ligand (L-asparagine). Clones that no longer respond to Asn but still respond to 5-ALA are identified as successful specificity-switch mutants.
- Biosensor Validation: The lead mutant (e.g., AC103-3H) is characterized. The dynamic range, sensitivity, and specificity of the biosensor are quantified by measuring fluorescence output in response to a concentration gradient of 5-ALA and other similar metabolites to rule out cross-reactivity.
- Application in High-Throughput Screening: The validated biosensor strain is used as a chassis for engineering metabolic pathways. A library of pathway variants is generated and spread on plates. Colonies with higher 5-ALA production are identified by their more intense red color, enabling rapid visual screening of millions of clones [51].
Genetically Encoded Biosensor Development
Instrument Deep-Dive: BD FACS Systems Comparison
For researchers considering FACS, the BD Biosystems portfolio offers different tiers of performance. The BD FACSCalibur is a legacy analyzer now discontinued [53]. The following table compares two modern high-performance sorters.
| Specification | BD FACSAria Fusion [48] [52] | BD FACSAria III [49] |
|---|---|---|
| Max Lasers | Configurable, upgradable (e.g., 405, 488, 640nm base) [52] | Up to 5 lasers, expandable via field upgrade [49] |
| Max Colors | Up to 14 colors (in a 3-laser configuration) [52] | Up to 18 colors simultaneously [49] |
| Optical System | Fixed-alignment, gel-coupled cuvette flow cell; Octagon & Trigon detection [48] | Fixed-alignment, gel-coupled cuvette flow cell; Patented Octagon & Trigon detection [49] |
| Integrated Biosafety | Standard (Class II Type A2 Biosafety Cabinet & Aerosol Management System) [48] | Not standard (relies on external cabinet) |
| Key Differentiating Feature | Fully integrated biosafety for operator and sample protection, ideal for CL2 work [48] [52] | Proven dependability and flexibility for a wide range of research applications [49] |
| Sort Collection | Tubes, slides, plates (with cooling/heating) [52] | Tubes, slides, plates |
Case Study: Directed Evolution of a 5-ALA Biosensor
This case exemplifies the power of biosensors in a directed evolution context. The goal was to create a biosensor for 5-aminolevulinic acid (5-ALA) to screen for high-producing engineered microbial strains [51].
- Challenge: No natural transcription factor for 5-ALA was known, preventing direct biosensor construction.
- Directed Evolution Solution:
- Scaffold Selection: The L-asparagine-responsive TF AsnC was chosen due to the structural similarity between Asn and 5-ALA.
- Diversification: A library of AsnC mutants was created via saturation mutagenesis of key amino acid sites.
- Selection/Screening: A high-throughput positive-negative screening was employed to find mutants that lost response to Asn but gained response to 5-ALA.
- Outcome: A mutant TF, AC103-3H, was isolated. When used to control RFP expression, it created a functional whole-cell biosensor (strain EAC103-3H). This biosensor successfully correlated fluorescence intensity to 5-ALA concentration, enabling the rapid visual screening of thousands of E. coli colonies on agar plates to identify top producers, a task that is prohibitively slow with traditional methods like HPLC or Ehrlich's reagent [51].
The Scientist's Toolkit: Essential Research Reagents and Materials
The following table lists key reagents and materials essential for implementing the described HTS technologies.
| Item | Function/Application | Example/Specification |
|---|---|---|
| Fluorochrome-conjugated Antibodies | Tag specific cell surface or intracellular markers for detection and sorting by FACS. | Anti-human FoxP3 antibody conjugated to BD Horizon V450 Dye [49]. |
| n-Dodecyl-β-D-Maltoside (DDM) | Mass spectrometry-compatible, non-ionic detergent for cell lysis. Improves recovery of membrane proteins in proteomic sample prep for assays like PISA [54]. | Used at 0.2% concentration [54]. |
| Phage Display Library | A diverse collection of filamentous phage clones, each displaying a unique peptide or protein fragment for screening against a target. | Used to identify novel biomarkers and therapeutic ligands for gastric cancer [50]. |
| 5-Aminolevulinic Acid (5-ALA) | Target metabolite. A non-protein amino acid and precursor for porphyrin compounds, used in photodynamic therapy and as a biofertilizer [51]. | Serves as the inducer molecule for the engineered biosensor [51]. |
| L-Asparagine (Asn) | Native ligand for the wild-type AsnC transcription factor. Used in negative selection during biosensor engineering to eliminate clones that did not switch specificity [51]. | Used to confirm loss of original TF function [51]. |
| Sorting Nozzles | Interchangeable tips that shape the sample stream into droplets for sorting. Different sizes accommodate different cell types. | A choice of nozzles for sorting a wide range of cell sizes on the BD FACSAria Fusion [52]. |
| Rp-8-Br-cGMPS (sodium salt) | Rp-8-Br-cGMPS (sodium salt), MF:C10H10BrN5NaO6PS, MW:462.15 g/mol | Chemical Reagent |
| p-NH2-Bn-oxo-DO3A | p-NH2-Bn-oxo-DO3A|Bifunctional Chelator |
The optimization of enzymes for industrial biocatalysis represents a cornerstone of sustainable manufacturing, particularly in the pharmaceutical sector where demands for precision, efficiency, and greener processes continue to intensify. This landscape is primarily shaped by two complementary approaches: trait-based optimization, which leverages natural enzyme diversity and predefined functional characteristics, and directed evolution, which employs iterative rounds of mutation and selection to engineer improved biocatalysts. While trait-based methods benefit from nature's evolutionary wisdom, directed evolution actively mimics and accelerates these natural processes in laboratory settings to achieve performance metrics beyond natural limits [55] [56].
Industrial applications, especially in drug development and manufacturing, require enzymes that not only exhibit high catalytic activity but also maintain robust stability under process conditions that often include non-aqueous solvents, elevated temperatures, and extreme pH levels. The fundamental challenge in biocatalyst engineering lies in the frequent activity-stability tradeoff, where enhancing one property often compromises the other due to the intricate balance between structural flexibility and rigidity required for enzymatic function [57] [58]. This case study examines contemporary strategies for evolving enzyme stability and activity, comparing their methodological frameworks, performance outcomes, and applicability to industrial biocatalysis.
Directed Evolution: Methodological Framework and Workflows
Directed evolution has emerged as a powerful methodology for engineering biocatalysts without requiring comprehensive prior knowledge of enzyme structure-function relationships. This approach mimics natural evolution through iterative cycles of diversity generation, screening, and variant selection to progressively enhance desired enzymatic properties [55].
Library Creation and Diversity Generation
The initial critical step involves creating genetic diversity within a parent enzyme sequence. While traditional methods like error-prone PCR and DNA shuffling remain widely used, recent advances have focused on designing "smarter" libraries that restrict sequence space to regions most likely to yield improvements [55]. Key methodological advancements include:
- Iterative Saturation Mutagenesis (ISM): Systematically targets specific residues for saturation mutagenesis to explore combinatorial mutations while minimizing library size [55].
- Site-Saturating Mutagenesis: Comprehensive substitution of single amino acid positions to assess their individual contributions to enzyme function.
- Hybrid Approaches: Combining random mutagenesis with structure-guided rational design to balance exploration and precision in library design [55].
The emergence of computational tools has significantly enhanced library design by identifying "key beneficial mutations" and predicting their potential impacts on protein folding and activity [55].
High-Throughput Screening and Selection Technologies
Identifying improved variants from vast mutant libraries represents the most significant bottleneck in directed evolution. Recent technological innovations have dramatically increased screening throughput and accuracy:
- Fluorescence-Activated Cell Sorting (FACS): Enables ultrahigh-throughput screening of up to 10^8 variants per day by coupling enzyme activity to fluorescent signals [55].
- Microfluidic Systems: Drop-based microfluidics confining substrate and single cells displaying enzyme variants in aqueous droplets for efficient screening [55].
- Phage-Assisted Continuous Evolution (PACE): Utilizes a fixed-volume vessel where phage replication is linked to target enzyme function, enabling continuous evolution without manual intervention [55].
- Enzyme Proximity Sequencing (EP-Seq): A novel deep mutational scanning method that leverages peroxidase-mediated radical labeling with single-cell fidelity to simultaneously assess thousands of mutations for both stability and catalytic activity [57].
These advanced screening platforms have dramatically reduced the time and resource requirements for directed evolution campaigns, with some pharmaceutical companies aiming to complete rounds of directed evolution within 7-14 days [59].
Experimental Platforms and Performance Metrics
Quantitative Comparison of Evolution Platforms
The table below summarizes the key characteristics and performance metrics of major directed evolution platforms:
Table 1: Performance Comparison of Directed Evolution Platforms
| Platform | Throughput (Variants) | Timeframe | Key Advantages | Primary Applications |
|---|---|---|---|---|
| FACS-based Screening | 10^8 per day | Days to weeks | Ultrahigh-throughput, high sensitivity | Enzyme activity, surface display systems [55] |
| Microfluidic Droplets | 10^8 per day | Hours to days | Minimal reagent use (150 μL for 10^8 variants), compartmentalization | Single-enzyme kinetics, pathway engineering [55] |
| PACE System | Continuous | Days | Autonomous evolution without intervention, direct genotype-phenotype linkage | Evolution of novel specificities, continuous improvement [55] |
| EP-Seq | 6,399+ mutants in parallel | Single experiment | Simultaneous stability & activity measurement, detailed fitness landscapes | Comprehensive mutational analysis, tradeoff studies [57] |
| Automated In Vivo Engineering | Limited by host capacity | Weeks | Growth-coupled selection, integrated hypermutation systems | Metabolic pathway engineering, in vivo optimization [60] |
Key Performance Metrics for Industrial Biocatalysis
Evaluating biocatalyst performance requires multiple metrics to assess industrial viability:
- Total Turnover Number (TTN): Defines the total number of catalytic cycles an enzyme completes before deactivation, with industrial processes often requiring values exceeding 10,000-50,000 [58].
- Product Concentration: Achievable product concentration directly impacts downstream processing costs, with high concentrations (>100 g/L) being economically essential [58].
- Volumetric Productivity: Measures the product formed per unit volume per time (gproduct L^{-1} h^{-1}), critical for determining reactor size and capital costs [60].
- Operational Stability: Enzyme performance under process conditions, typically measured as half-life or duration of maintained activity, often more relevant than thermodynamic stability [58].
- Space-Time Yield: The amount of product formed per unit reactor volume per time, integrating both catalytic efficiency and process intensification [59].
Case Study: D-Amino Acid Oxidase Engineering via EP-Seq
Experimental Workflow and Methodology
A recent breakthrough in understanding activity-stability tradeoffs was achieved through Enzyme Proximity Sequencing (EP-Seq) applied to D-amino acid oxidase (DAOx) from Rhodotorula gracilis [57]. The experimental methodology encompassed:
Library Construction:
- Site saturation mutagenesis across the entire DAOx coding region
- Incorporation of 15-nucleotide unique molecular identifiers (UMIs) for variant tracking
- Yeast surface display system with Aga2p anchoring for cell-surface expression
Stability/Expression Assessment:
- Induced expression (48 h, 20°C, pH 7) of variant library
- C-terminal His-tag staining with primary and fluorescent secondary antibodies
- FACS sorting into four bins based on expression level
- Expression fitness score calculation: ( {\log}2({\beta}{v}/{\beta}{wt}) ), where ( {\beta}{v} ) and ( {\beta}_{wt} ) represent variant and wild-type expression scores [57]
Activity Profiling:
- HRP-mediated phenoxyl radical coupling reaction at yeast surface
- Tyramide-488 labeling intensity correlated with enzymatic activity
- FACS sorting into four activity bins
- Activity fitness score calculation analogous to expression scoring
- High reproducibility (Pearson's r = 0.96 for activity replicates) [57]
Diagram 1: EP-Seq workflow for comprehensive variant analysis.
Key Findings and Quantitative Results
The EP-Seq analysis of 6,399 missense mutations in DAOx revealed critical insights into activity-stability relationships:
Table 2: Quantitative Results from DAOx Deep Mutational Scanning
| Parameter | Wild-Type DAOx | Improved Variants | Measurement Technique |
|---|---|---|---|
| Expression Fitness Distribution | Reference (0) | -2.5 to +1.5 log2 scale | FACS + NGS quantification [57] |
| Activity Fitness Distribution | Reference (0) | -3.0 to +2.0 log2 scale | Proximity labeling + FACS [57] |
| Activity-Stability Correlation | Baseline | R = 0.68 (positive correlation) | Linear regression of fitness scores [57] |
| Reproducibility (Activity) | N/A | Pearson's r = 0.96 | Biological replicates [57] |
| Reproducibility (Expression) | N/A | Pearson's r = 0.94 | Biological replicates [57] |
| Hotspot Identification | Active site | Distal regions affecting allostery | Fitness landscape mapping [57] |
The study demonstrated that catalytic activity constrains folding stability during natural evolution, identifying specific "hotspots" distant from the active site as candidates for mutations that improve catalytic activity without sacrificing stability [57]. This finding challenges simplistic tradeoff models and reveals opportunities for simultaneous optimization of both properties through targeted engineering of allosteric networks.
Emerging Technologies: AI and Automation in Biocatalysis
Machine Learning-Guided Enzyme Engineering
Artificial intelligence and machine learning are rapidly transforming enzyme engineering paradigms:
- Protein Language Models: Tools like ProtT5, Ankh, and ESM2 leverage unlabeled sequence data to predict functional relationships and guide zero-shot predictions without experimental data [61].
- Fitness Landscape Navigation: ML models trained on experimental data can predict mutation effects, prioritizing beneficial combinations and epistatic interactions [61].
- De Novo Enzyme Design: Generative models including RFdiffusion enable backbone design and active site scaffolding for novel enzyme functions [60].
Industry applications demonstrate that ML-guided directed evolution can successfully optimize challenging enzymes such as halogenases for late-stage functionalization of complex macrolides and ketoreductases for manufacturing cancer drug precursors [61].
Automated and Continuous Evolution Platforms
Integrated automated workflows represent the cutting edge of biocatalyst development:
- Automated Biofoundries: Robotic systems enable high-throughput implementation of Design-Build-Test-Learn cycles, significantly accelerating engineering timelines [60].
- Growth-Coupled Selection: Linking enzyme function to microbial fitness enables autonomous enrichment of improved variants without manual screening [60].
- In Vivo Hypermutation Systems: Targeted increase of mutation rates in specific genomic regions combined with continuous cultivation platforms [60].
These integrated systems are particularly valuable for optimizing multi-enzyme pathways and complex metabolic engineering tasks where traditional approaches face significant scalability challenges.
Research Reagent Solutions for Enzyme Engineering
Table 3: Essential Research Reagents and Platforms for Directed Evolution
| Reagent/Platform | Function | Key Applications | Examples/References |
|---|---|---|---|
| Yeast Surface Display | Eukaryotic expression and display system | Enzyme stability assessment, eukaryotic folding requirements | Aga2p fusion system for DAOx [57] |
| Fluorescence-Activated Cell Sorter (FACS) | High-throughput cell sorting based on fluorescence | Ultrahigh-throughput screening of enzyme libraries [55] | Sorting 10^8 variants per day [55] |
| Tyramide-Based Proximity Labeling | Enzyme activity-dependent cell surface labeling | Massively parallel activity screening | EP-Seq for oxidase activity [57] |
| Error-Prone PCR Kits | Introduction of random mutations throughout gene | Library generation for unexplored sequence spaces | Commercial kits from suppliers like NEB [55] |
| Site-Saturation Mutagenesis Kits | Targeted substitution of specific codons | Focused library design for known hotspots | Commercial kits from suppliers like ThermoFisher [55] |
| Microfluidic Droplet Generators | Compartmentalization of single cells/enzymes | Single-cell enzyme assays, minimal reagent use | Drop-based microfluidics [55] |
| Phage Display Vectors | In vitro selection platform linking genotype to phenotype | Protein-ligand interactions, binding affinity maturation | PACE system development [55] |
Comparative Analysis: Trait-Based Versus Directed Evolution Approaches
The choice between trait-based and directed evolution strategies depends on multiple factors, including starting enzyme characteristics, desired improvements, and available resources:
Performance and Applicability Comparison
Table 4: Trait-Based vs. Directed Evolution Approach Comparison
| Parameter | Trait-Based Approach | Directed Evolution |
|---|---|---|
| Starting Point | Natural enzyme diversity, metagenomic libraries [59] | Single parent enzyme with some target function [55] |
| Required Prior Knowledge | High (sequence-function relationships) | Low to moderate [55] |
| Library Size | Large (searching natural diversity) | Focused (10^3-10^8 variants) [55] |
| Throughput Potential | Moderate (dependent on screening assay) | High (FACS: 10^8/day) [55] |
| Development Timeline | Weeks to months | Days to weeks (PACE: days) [55] [59] |
| Automation Potential | Moderate | High (integrated automated workflows) [60] |
| Success Rate | Variable (dependent on natural diversity) | High (iterative improvement) [55] |
| Optimal Application Scope | Novel function discovery, metagenomic mining [59] | Optimizing existing activities, stability engineering [55] |
Integrated Workflow for Community Optimization
The most effective biocatalyst development strategies increasingly combine elements from both approaches:
Diagram 2: Integrated workflow combining trait-based and directed evolution approaches.
The evolution of enzyme stability and activity for industrial biocatalysis has progressed from simple trait selection to sophisticated integrated engineering platforms. Directed evolution excels at optimizing specific enzyme properties, while trait-based approaches leverage nature's diversity for novel function discovery. The emerging paradigm combines both strategies with AI-guided design and automated implementation, dramatically accelerating development timelines from months to weeks or even days [59] [60].
Future advancements will likely focus on overcoming the persistent activity-stability tradeoff through deeper understanding of allosteric networks and dynamic structural relationships [57]. As the field matures, standardized performance metrics and benchmarking protocols will become increasingly important for comparing biocatalysts across different studies and applications [58]. The integration of machine learning, automated workflows, and high-throughput experimental platforms promises to further accelerate the development of robust, efficient biocatalysts for sustainable pharmaceutical manufacturing and industrial chemistry.
Synthetic Directed Evolution (SDE) for Engineering Complex Plant and Microbial Traits
The optimization of biological systems for agriculture and medicine has long relied on two contrasting paradigms: trait-based engineering and directed evolution. Trait-based engineering operates on a rational design principle, where specific, known traits or genes are deliberately introduced or modified based on prior knowledge of structure and function [32]. Conversely, synthetic directed evolution (SDE) mimics natural selection in an accelerated form, employing iterative cycles of artificial diversification and selection to evolve genes or pathways toward a desired function without requiring comprehensive prior knowledge [31]. This guide provides a comparative analysis of modern SDE methodologies, offering experimental protocols and reagent solutions to empower research in this rapidly advancing field.
Core Principles and Workflow of SDE
SDE operates through a cyclic process of diversification and selection [32] [31]. In the diversification stage, researchers introduce mutations into specific gene sequences, generating a library of gene variants. In the subsequent selection stage, the host population is subjected to a specific pressure to identify individuals with the desired traits. Selected variants can undergo multiple rounds of this process to further enrich and amplify the traits of interest [32].
This process can be imagined as navigation across high-dimensional "fitness landscapes," which map each genetic sequence to a measure of fitness. The goal of directed evolution is to find the highest peaks on this landscape [47]. The following diagram illustrates the core SDE workflow and its relationship to the fitness landscape.
Comparative Analysis of SDE Diversification Methods
Various strategies exist for the diversification stage of SDE, each with distinct mechanisms, advantages, and applications. The table below summarizes the performance and characteristics of key methods.
Table 1: Comparison of SDE Diversification Methods and Applications
| Method | Key Mechanism | Primary Application Context | Advantages | Limitations & Key Experimental Outcomes |
|---|---|---|---|---|
| Error-Prone PCR [32] | Random mutations via low-fidelity PCR. | In vitro protein engineering. | Simple, rapid, no structural knowledge needed. | Limited mutation rate, restricted amino acid variants, adjacent mutations unlikely. |
| DNA Shuffling [32] | Recombination of larger gene fragments from homologous genes. | In vitro evolution of multi-gene pathways. | Exchanges functional domains, can integrate beneficial mutations. | Requires sequence homology, can be labor-intensive. |
| CRISPR-Cas NHEJ [31] | Non-homologous end joining repair of Cas-induced double-strand breaks. | Targeted gene disruption in plants. | High efficiency in plants, enables gene knockouts. | Introduces unpredictable indels; used to evolve herbicide-resistant rice SF3B1 [31]. |
| Base Editing [31] | Cas nickase fused to deaminase for precise point mutations. | Protein optimization in plants. | No double-strand breaks, predictable C->T or A->G transitions. | Limited to specific base changes; generated herbicide-resistant rice ACCase [31]. |
| EvolvR [31] | Cas nickase fused to error-prone polymerase for continuous diversification. | In vivo continuous evolution in microbes. | Targeted, user-defined mutation window. | Primarily demonstrated in prokaryotic systems. |
| MAGE [32] | Multiplexed recombineering with oligonucleotides. | Multiplex genome engineering in microbes. | Enables simultaneous diversification of multiple genomic sites. | Primarily effective in microbial systems like E. coli. |
Detailed Experimental Protocols for Key SDE Studies
Protocol 1: Engineering Herbicide Resistance in Rice via CRISPR-Cas9
This protocol is adapted from a study that evolved herbicide-resistant variants of the rice splicing factor SF3B1 [31].
- Library Design and Construction: Design a tiled library of single-guide RNAs (sgRNAs) covering the entire coding sequence of the target gene. Clone the sgRNA library into an appropriate CRISPR-Cas9 vector.
- Plant Transformation: Introduce the sgRNA-Cas9 construct into rice cells using Agrobacterium tumefaciens-mediated transformation.
- Selection and Screening: Culture the transformed plant tissue on a medium containing the target herbicide. Select the surviving plantlets that show resistance.
- Genotypic Validation: Sequence the target gene in resistant plants to identify the specific mutations conferring the resistance.
- Phenotypic Confirmation: Regenerate the mutant plants and test their resistance to the herbicide and ensure the normal splicing function is retained.
Protocol 2: Continuous Evolution in Microbes Using Targeted Mutagenesis
This protocol outlines the use of tools like EvolvR for in vivo directed evolution in bacterial systems [31] [47].
- System Assembly: Construct a plasmid expressing the EvolvR system, which consists of a nickase Cas9 (nCas9, D10A) fused to an error-prone DNA polymerase and a gene-specific sgRNA.
- Library Generation: Transform the assembled plasmid into the host microbial strain. The nCas9 induces a single-strand nick in the target DNA, and the tethered error-prone polymerase introduces mutations during repair, generating a diverse variant library in vivo.
- Selection Pressure: Apply the desired selective pressure to the culture. This could involve adding an antibiotic, toxin, or requiring the utilization of a novel substrate.
- Variant Isolation: Isolate surviving clones, which harbor beneficial mutations. This can be done through simple plating or using advanced sorting methods like FACS or microfluidics [47].
- Iteration and Analysis: Use the selected clones as the starting point for subsequent rounds of evolution or sequence them to identify the causative mutations.
Visualization of Key SDE Experimental Workflows
The following diagram illustrates the specific workflow for evolving herbicide resistance in plants, a key application of SDE.
The Scientist's Toolkit: Essential Research Reagents
Successful implementation of SDE requires a suite of specialized reagents and tools. The table below details essential components for building an SDE pipeline.
Table 2: Key Research Reagent Solutions for SDE
| Reagent/Tool | Function in SDE | Specific Examples & Notes |
|---|---|---|
| Diversification Tools | Creates genetic diversity at the target locus. | CRISPR-Cas9 (for NHEJ), Base Editors (for point mutations), EvolvR (for continuous evolution), error-prone PCR kits [32] [31]. |
| Vector Systems | Delivers genetic components into the host. | Plant transformation vectors (e.g., for Agrobacterium), microbial expression plasmids, viral delivery systems. |
| Selection Agents | Applies selective pressure to enrich for desired traits. | Herbicides (e.g., for evolving resistance in crops), antibiotics, novel carbon sources [31]. |
| sgRNA Library | Guides nucleases to specific genomic loci for diversification. | A pooled library of sgRNAs tiling a gene of interest; essential for targeted CRISPR-SDE [31]. |
| Cell Sorting/Microfluidics | Enables high-throughput screening and selection of variants. | Fluorescence-Activated Cell Sorting (FACS) or emerging microfluidic platforms that allow selection based on dynamic phenotypes [47]. |
| Machine Learning Models | Acts as a surrogate to predict fitness, guiding exploration. | Gaussian Processes or other models used in Bayesian Optimization to balance exploration and exploitation [62]. |
| Spphpspafspafdnlyywdq | HER2/neu Multi-Epitope Peptide | SPPHPSPAFSPAFDNLYYWDQ is a multi-epitope class II rat HER2/neu peptide for cancer vaccine research. For Research Use Only. Not for human use. |
| Peli1-IN-1 | Peli1-IN-1, MF:C20H16O4, MW:320.3 g/mol | Chemical Reagent |
Synthetic directed evolution represents a powerful, selection-driven alternative to purely rational, trait-based design. As the showcased protocols and comparisons demonstrate, SDE technologies like CRISPR-based base editing and in vivo mutagenesis systems enable the rapid evolution of complex traits in both plants and microbes. The integration of advanced screening methods and machine learning is further refining SDE strategies, optimizing the navigation of fitness landscapes without the absolute requirement for sequencing [47] [62]. This synergy between experimental evolution and computational prediction is unlocking new frontiers in engineering biology for agricultural resilience and drug development.
Overcoming Challenges: Managing Variability and Optimizing Outcomes
In the pursuit of robust and productive bioprocesses, scientists confront a fundamental challenge: how to effectively optimize biological systems despite complex, often opaque, sequence-to-function relationships. This challenge frames a critical methodological divide between trait-based approaches, which aim to control and fine-tune existing process parameters, and directed evolution approaches, which engineer the biological components themselves for enhanced performance. Trait-based optimization operates on the principle that extracellular conditions—such as nutrient feed, temperature, and pH—can be dynamically controlled to maximize yield from a given biological host. In contrast, directed evolution seeks to improve the host's intrinsic capabilities through iterative mutagenesis and selection, effectively altering its genetic makeup to achieve the desired trait. This guide provides a structured comparison of these paradigms, offering a methodical framework for diagnosing yield variations and selecting the appropriate optimization strategy based on specific research goals and constraints. Yield variations in bioprocessing present a multi-faceted problem, often stemming from inconsistencies in upstream cell culture conditions, raw material variability, uncalibrated sensors, or the inherent biological complexity of the production host [63] [64] [65]. A systematic approach is therefore essential for identifying root causes and implementing effective corrective actions.
Trait-Based Optimization: Controlling the Process
Trait-based optimization focuses on maximizing yield by tightly controlling the bioprocess environment and operating parameters. This approach is particularly valuable when working with a genetically fixed production host.
Core Principles and Data-Driven Methodologies
The foundation of trait-based optimization lies in the precise monitoring and control of Critical Process Parameters (CPPs) to influence key yield indicators. A recent industrial case study on monoclonal antibody (mAb) production exemplifies this data-driven approach. Researchers applied machine learning (ML) regression models to historical batch records to predict three critical yield indicators: Bioreactor Final Weight (BFW), Harvest Titer (HT), and Packed Cell Volume (PCV) [63]. Their methodology, outlined below, provides a robust protocol for modern bioprocess analysis.
Table: Machine Learning Model Performance for Yield Prediction
| Yield Indicator | Best-Performing Model | Reported Performance (R²) | Modeling Difficulty |
|---|---|---|---|
| Bioreactor Final Weight (BFW) | Support Vector Regression (SVR) | 0.978 | Accurately predictable |
| Harvest Titer (HT) | Various Models (Random Forest, Gradient Boosting) | Difficult to model accurately | High |
| Packed Cell Volume (PCV) | Various Models (Random Forest, Gradient Boosting) | Difficult to model accurately | High |
Experimental Protocol: Data-Driven Yield Analysis [63]
- Data Collection & Preprocessing: Gather historical batch records encompassing process inputs, monitored variables, and batch outcomes. Exclude batches with missing critical values. Perform outlier detection through visualization tools (e.g., histograms, scatter plots) and normalize numerical features to ensure comparable scales.
- Exploratory Data Analysis (EDA): Use descriptive statistics and correlation analysis to understand relationships between process parameters and yield outcomes. Generate heatmaps to visualize correlations and identify candidate features for modeling.
- Machine Learning Model Development: Train multiple regression models (e.g., Random Forest, Gradient Boosting, SVR) to predict key yield indicators. Evaluate model performance to identify the most influential process parameters through sensitivity analysis.
- Process Optimization Exploration: Apply optimization algorithms like Sequential Least-Squares Programming (SLSQP) to suggest parameter combinations associated with improved yield estimates compared to historical averages.
For fed-batch processes, model-based frameworks like OptFed have been developed to address sub-optimal feed profiles. This methodology uses measurements of bioreactor volume, biomass, and product to fit kinetic constants and solve an optimal control problem, dynamically determining the best feed rate and temperature to maximize metrics like the product-to-biomass yield [66].
The Scientist's Toolkit: Essential Reagents and Solutions
Table: Key Research Reagent Solutions for Bioprocess Monitoring and Control
| Item | Function/Benefit |
|---|---|
| Process Analytical Technology (PAT) | Enables real-time monitoring of CPPs (e.g., pH, DO, cell density, nutrient levels) for automated, in-process control [64]. |
| Auto Calibrate Software (e.g., BioFlo systems) | Automates sensor calibration (e.g., for DO sensors) to ensure high-precision readings and reduce batch-to-batch variability [64]. |
| Statistical Process Control (SPC) Charts | A statistical tool to detect patterns, trends, or outliers in process data, helping to identify variations outside the acceptable range [65]. |
Figure: Trait-Based Optimization Workflow
Directed Evolution: Engineering the Biological Agent
Directed evolution mimics natural selection to optimize proteins or entire metabolic pathways without requiring prior mechanistic knowledge of the system. It is a powerful alternative when process optimization alone is insufficient.
Modern Methodologies and Machine Learning Enhancement
The classical directed evolution cycle involves iterative rounds of mutagenesis to create genetic diversity and selection to isolate improved variants. However, standard "greedy" selection of top-performing variants each generation is prone to trapping in local optima on rugged fitness landscapes [20] [3]. Emerging strategies aim to improve the efficiency of this navigation.
Experimental Protocol: Standard Directed Evolution [3]
- Library Generation (Mutagenesis): Create a diverse library of gene variants. Common methods include:
- Error-prone PCR: Introduces random point mutations across the entire gene.
- DNA Shuffling: Recombines genes from different parents to create chimeric variants.
- Site-Saturation Mutagenesis: Targets specific residues for exhaustive mutagenesis.
- Selection or Screening: Isolate variants with desired traits. This can be high-throughput (e.g., using FACS or display techniques) or lower-throughput (e.g., colorimetric assays on plates) depending on the trait and resources.
- Characterization & Iteration: Characterize the selected hits and use them as templates for subsequent rounds of evolution.
To overcome the limitations of simple directed evolution, Active Learning-assisted Directed Evolution (ALDE) has been developed. ALDE integrates machine learning with wet-lab experimentation to navigate epistatic landscapes more efficiently. In a case study optimizing a non-native cyclopropanation reaction in a protoglobin, ALDE improved the product yield from 12% to 93% in just three rounds by effectively modeling the epistatic interactions between five active-site residues [67].
Another advanced strategy involves using parameterized "selection functions" that tune the balance between exploration (searching new areas of sequence space) and exploitation (selecting known high-fitness variants). Simulations on empirical fitness landscapes show that such alternative strategies can lead to up to a 19-fold increase in the probability of finding the global fitness peak compared to standard methods [20].
Table: Comparison of Directed Evolution Techniques
| Technique | Key Principle | Advantage | Disadvantage |
|---|---|---|---|
| Standard DE | Iterative greedy selection of top variants | Simple, established workflow | Prone to local optima, inefficient on rugged landscapes [20] |
| ALDE | ML batch Bayesian optimization guides library selection | Efficiently handles epistasis; high success in few rounds [67] | Requires computational infrastructure and expertise |
| Tuned Selection Functions | Probabilistic selection to balance exploration/exploitation | Can increase probability of finding global optimum [20] | Requires simulation or prior landscape knowledge |
Figure: Directed Evolution with Active Learning
Comparative Analysis and Decision Framework
The choice between trait-based and directed evolution strategies is not mutually exclusive, but the initial focus depends on the specific nature of the yield limitation.
Table: Strategy Comparison: Trait-Based vs. Directed Evolution
| Aspect | Trait-Based Optimization | Directed Evolution |
|---|---|---|
| Primary Focus | Optimizing extracellular process parameters (e.g., feed, pH, temperature) [63] [68] | Engineering the intrinsic capabilities of the biological agent (host cell or enzyme) [3] [67] |
| Knowledge Requirement | Relies on process data and understanding of CPPs [63] | Requires no prior mechanistic knowledge of sequence-to-function [20] |
| Typical Timeframe | Short-to-medium term (process control adjustments) | Medium-to-long term (multiple iterative cycles) |
| Key Tools | PAT, ML, DoE, Statistical Process Control [63] [65] | Mutagenesis methods, FACS, display techniques, Active Learning [3] [67] |
| Ideal Use Case | Solving issues of process consistency, batch variability, and sub-optimal control [64] [65] | Improving intrinsic properties like enzyme activity, stability, or substrate specificity [3] |
| Reported Outcome | 19% improvement in product-to-biomass yield via dynamic modeling [66] | 12% to 93% product yield improvement in an epistatic enzyme landscape [67] |
Integrated Troubleshooting Workflow
A methodical approach to yield variation involves sequential investigation:
- Audit the Process First: Before altering the biological host, exhaustively investigate process parameters. This includes reviewing batch records, using multivariate analysis to find parameter-yield correlations, ensuring raw material consistency, and verifying equipment calibration [65]. In many cases, the root cause lies here, and solutions are faster to implement.
- Resort to Directed Evolution for Intrinsic Limitations: If process optimization is maximized but yield remains limited by the host's innate capabilities—such as low specific productivity, poor thermostability, or undesired kinetics—directed evolution becomes the necessary path.
- Consider Hybrid Strategies: For the most challenging problems, combine both approaches. Use directed evolution to create a superior production host, then apply trait-based optimization to develop a finely tuned process that maximizes the potential of the evolved variant.
Navigating yield variations in bioprocesses requires a disciplined, diagnostic mindset. The dichotomy between trait-based optimization and directed evolution provides a clear framework for action. Trait-based strategies offer powerful, data-driven tools to bring a process under statistical control and maximize the output from a given biological system. Conversely, directed evolution, especially when enhanced with active learning, allows researchers to fundamentally redesign the biological system itself, breaking through performance plateaus imposed by intrinsic limitations. The most effective bioprocess engineers are those who can accurately diagnose the source of a yield problem and strategically deploy the most appropriate toolkit, whether it focuses on the process, the producer, or an integrated combination of both.
In ecology and bioprocessing, the conventional wisdom holds that greater species diversity inherently leads to greater functional diversity—the variety of ecological functions performed by organisms within a community. This premise underpins countless conservation strategies and biotechnological approaches to community engineering. However, emerging research reveals a troubling paradox: under specific conditions, increasing species diversity can actually decrease functional diversity, potentially compromising ecosystem stability and biotechnological yield. This paradox represents a critical challenge for researchers, scientists, and drug development professionals working with microbial communities, engineered ecosystems, and synthetic biology platforms.
The tension between species diversity and functional diversity arises from complex eco-evolutionary processes that shape trait distributions in competitive environments [2]. As species richness increases within a confined niche, intensified competition often drives evolutionary trait narrowing, forcing species to specialize on narrower resource spectra to avoid competitive exclusion. Consequently, while taxonomic metrics may suggest healthy biodiversity, the actual functional capacity of the community may be substantially eroded—a phenomenon with profound implications for ecosystem functioning, resilience, and biotechnological optimization [69] [2].
This comparative guide examines the paradox of diversity through the contrasting lenses of two predominant approaches for community optimization: trait-based approaches, which rely on rational design principles to assemble communities with prescribed functions, and directed evolution strategies, which harness evolutionary pressures to optimize community performance. By synthesizing recent findings from ecological modeling, experimental evolution, and synthetic biology, we provide researchers with a framework for navigating the complex relationship between species counts and functional capacity in engineered biological systems.
Quantitative Evidence: Documenting the Paradox Across Ecosystems
Empirical and theoretical studies across diverse ecosystems consistently demonstrate that species diversity does not reliably predict functional diversity. The following table summarizes key findings from foundational studies documenting this paradox.
Table 1: Empirical Evidence of the Diversity Paradox Across Ecosystems
| Study System | Species Diversity Trend | Functional Diversity Trend | Proposed Mechanism | Citation |
|---|---|---|---|---|
| Galápagos land snails | Higher in species-rich communities | Lower in species-rich communities | Evolutionary trait narrowing in competitive communities | [2] |
| Coral reefs under urban stress | Increasing (taxonomic & phylogenetic) | Decadal decline | Chronic urbanization stress filtering functional traits | [69] |
| Global plant communities (remote sensing) | Seasonal variation | Biome-specific seasonal dynamics, sometimes opposing | Phenological shifts and environmental filtering | [70] |
| Modeled trophic webs | Variable | Higher functional diversity increased resistance and resilience | Complementarity effects and functional redundancy | [71] |
| Urban avian assemblages | Variable | Not consistently different from non-urban; sometimes higher after richness correction | Habitat diversity within cities enables niche partitioning | [72] |
The consistency of these findings across disparate systems—from terrestrial snails to microbial communities—suggests the diversity paradox represents a fundamental ecological phenomenon rather than a system-specific anomaly. The Galápagos land snail study proved particularly illuminating, demonstrating that species in rich communities evolved narrower trait breadths to avoid competition, resulting in reduced overall trait space coverage despite increased species counts [2]. Similarly, coral reef monitoring documented a worrying decoupling between taxonomic and functional diversity metrics, with chronic urbanization stress systematically filtering out functional traits regardless of phylogenetic representation [69].
Experimental Approaches: Methodologies for Assessing the Paradox
Eco-Evolutionary Modeling Protocols
The theoretical foundation for understanding the diversity paradox relies on sophisticated eco-evolutionary models that integrate trait evolution with population dynamics:
Table 2: Experimental Protocols in Eco-Evolutionary Modeling
| Protocol Component | Implementation Details | Ecological Insight |
|---|---|---|
| Model framework | Quantitative genetic model tracking population density, trait means, and variances | Simultaneously captures ecological and evolutionary dynamics |
| Trait representation | Multiple trait dimensions with evolving covariance structures | Reveals how genetic correlations respond to selective pressures |
| Competition function | Resource competition based on trait similarity | Quantifies how niche overlap drives trait divergence |
| Evolutionary dynamics | Continuous-time adaptation of trait distributions | Shows how intraspecific variation responds to community context |
| Equilibrium criteria | Ecological and evolutionary equilibrium simultaneously | Identifies stable endpoints of community assembly |
The model follows a continuous-time framework where trait distributions evolve in response to competitive interactions. Each species is characterized by its population density (N), mean trait values (μ), and trait covariance matrix (G). The intrinsic growth rate of phenotypes is determined by their position in trait space, while competition arises from phenotypic similarity [2]. Implementation typically begins with randomly generated communities varying in initial species diversity, with analysis focusing on how functional diversity (measured as trait space coverage) changes as the community reaches eco-evolutionary equilibrium.
Trait-Based Assessment Methods
Empirical detection of the diversity paradox requires robust functional diversity metrics. The following experimental approaches are commonly employed:
Trait Measurement Protocols:
- Plant communities: Measurement of vegetative and regenerative traits for all species in sample plots, including leaf area, specific leaf area, plant height, and seed mass [73]
- Microbial communities: Genomic trait inference through KEGG pathway annotation combined with metatranscriptomic profiling to assess trait expression patterns [74]
- Remote sensing: Hyperspectral data analysis to estimate plant functional traits across landscapes and seasons [70]
Functional Diversity Metrics:
- Rao's Quadratic Entropy (Rao's Q): Combines species abundances and trait differences; effective at detecting both trait convergence (habitat filtering) and divergence (limiting similarity) [73]
- Functional Richness: Measures the range of trait values in a community; calculated via convex hull or kernel density estimation hypervolume approaches [70]
- Standardized Effect Sizes: Comparison of observed functional diversity to null model expectations to identify assembly processes [73]
Recent methodological evaluations have revealed significant limitations in existing functional diversity metrics, with many failing to satisfy basic mathematical requirements for ideal diversity measures [75]. Researchers should therefore employ multiple complementary metrics to obtain robust assessments of functional diversity.
Comparative Analysis: Trait-Based versus Directed Evolution Approaches
The diversity paradox presents distinct challenges and opportunities for the two predominant approaches to community optimization:
Table 3: Trait-Based versus Directed Evolution Approaches to Community Optimization
| Aspect | Trait-Based Approaches | Directed Evolution Approaches |
|---|---|---|
| Theoretical basis | Rational design based on known trait-function relationships | Artificial selection of high-performing communities |
| Implementation | Bottom-up assembly of consortia based on functional traits | Iterative propagation and selection of whole communities |
| Handling of diversity paradox | Explicit consideration of trait overlap and complementarity | Emergent resolution through selection on community function |
| Technical requirements | Detailed prior knowledge of species traits | High-throughput screening capabilities |
| Limitations | Incomplete trait knowledge; complex trait interactions | Historical contingency; limited predictability |
| Best applications | Systems with well-characterized component species | Complex communities with unknown structure-function relationships |
Trait-based approaches attempt to "solve the puzzle" of community assembly by carefully selecting species with complementary functional traits, analogous to how protein designers select amino acids based on their biochemical properties [9]. This approach succeeds when trait-function relationships are well-understood but struggles when emergent properties arise from unexpected species interactions.
Directed evolution approaches remain agnostic to the mechanistic basis of community function, instead selecting for overall performance through iterative cycles of propagation and variation [9]. This method harnesses evolutionary processes similar to those that create the diversity paradox, potentially leveraging them for biotechnological advantage rather than being constrained by them.
Visualization: Conceptual Framework and Experimental Workflows
Mechanistic Basis of the Diversity Paradox
Mechanistic Basis of the Diversity Paradox
This diagram illustrates the competing pathways through which increased species diversity affects functional diversity. The red pathway dominates under strong competition, driving evolutionary trait narrowing and creating the diversity paradox [2]. The green pathway represents the conventional expectation of enhanced niche partitioning, which prevails when competition is sufficiently weak or resource diversity sufficiently high to allow coexistence without trait contraction [76].
Experimental Workflow for Paradox Investigation
Experimental Workflow for Paradox Investigation
This workflow outlines the integrated experimental approach needed to investigate the diversity paradox, combining initial community assembly with simultaneous ecological and evolutionary monitoring [2] [74]. The critical innovation lies in tracking both trait means and variances across generations, as declining intraspecific variation drives the paradox despite potential stability in species-level trait averages.
Table 4: Essential Research Tools for Investigating Functional Diversity Relationships
| Tool/Resource | Application Context | Key Functionality | Implementation Considerations |
|---|---|---|---|
| TbasCO | Trait-based comparative 'omics | Identifies expression-based attributes of predefined traits | Requires time-series metatranscriptomics data; KEGG pathway database as trait library [74] |
| Rao's Q Calculator | Community ecology | Computes functional diversity using abundance-weighted trait differences | Sensitive to trait selection; less correlated with species richness than other metrics [73] |
| Hyperspectral Trait Mapping | Landscape ecology | Estimates plant functional traits through remote sensing | Enables large-scale monitoring; limited to optically-detectable traits [70] |
| Eco-Evolutionary Models | Theoretical ecology | Simulates simultaneous ecological and evolutionary dynamics | Requires quantitative genetic parameters; computationally intensive for diverse communities [2] |
| KEGG Pathway Database | Microbial ecology | Provides curated metabolic pathways for trait inference | Enzyme commission-based annotations may not capture regulatory differences [74] |
The paradox of diversity—where more species leads to less functional diversity—represents a fundamental challenge to conventional approaches in conservation biology, ecosystem management, and microbial community engineering. The evidence compiled in this guide demonstrates that the relationship between species diversity and functional diversity is complex, context-dependent, and powerfully shaped by eco-evolutionary dynamics.
For researchers and biotechnology professionals, these findings highlight the limitations of species-centric approaches to biodiversity conservation and community engineering. Successful navigation of the diversity paradox requires increased attention to functional trait distributions—including both interspecific and intraspecific variation—rather than simple taxonomic inventories. Specifically, we recommend:
- Adopting multidimensional assessments that simultaneously track taxonomic, phylogenetic, and functional diversity metrics
- Monitoring trait variances in addition to means, as declining intraspecific variation often drives the diversity paradox
- Considering temporal dynamics, as functional diversity exhibits meaningful seasonality in many systems [70]
- Balancing design approaches by combining trait-based rational design with directed evolution to harness both mechanistic understanding and emergent optimization
Ultimately, recognizing the diversity paradox does not diminish the importance of species conservation or diverse community assembly, but rather refines our understanding of how biodiversity translates into ecological function. By moving beyond species counts to embrace the complex interplay of traits, niches, and evolutionary dynamics, researchers can develop more effective strategies for managing and engineering biological communities in both natural and biotechnology contexts.
The pharmaceutical industry is undergoing a significant paradigm shift from traditional batch processing toward continuous manufacturing, driven by the need for enhanced reproducibility, efficiency, and product quality. This transition is supported by three interconnected technological pillars: Process Analytical Technology (PAT) for real-time monitoring, Model Predictive Control (MPC) for advanced process control, and automation systems for integrated execution. Regulatory agencies, including the U.S. Food and Drug Administration (FDA), have championed this evolution through initiatives like Quality by Design (QbD), which emphasizes proactive, science-driven methodologies over traditional reactive quality testing [77]. Within this technological context, a fundamental scientific question emerges: how can we best optimize complex manufacturing systems? This article frames the discussion around two competing approaches—trait-based optimization, which systematically controls known critical parameters, versus directed evolution, which iteratively selects for desirable outcomes without requiring complete mechanistic understanding. By comparing advanced process control methodologies through this conceptual lens, this guide provides researchers and drug development professionals with a framework for selecting and implementing strategies that enhance reproducibility in pharmaceutical manufacturing.
Theoretical Framework: Trait-Based versus Directed Evolution Approaches
The optimization of complex systems, whether biological or manufacturing, can be approached through two distinct philosophies, each with characteristic methodologies, advantages, and applications.
Trait-Based Approaches
Trait-based approaches operate on a fundamental principle: system performance can be optimized through systematic identification and control of critical traits or parameters. In pharmaceutical manufacturing, this is embodied by the QbD framework, where Critical Quality Attributes (CQAs) are prospectively defined, and Critical Process Parameters (CPPs) are controlled to ensure final product quality [77]. This methodology requires deep process understanding and relies on mechanistic models that establish cause-effect relationships between process parameters and product quality.
- Key Methodology: The implementation follows a defined workflow: (1) Define Quality Target Product Profile (QTPP), (2) Identify CQAs, (3) Perform risk assessment, (4) Design of Experiments (DoE) to establish parameter relationships, (5) Establish a design space, (6) Develop control strategy, and (7) Implement continuous improvement [77].
- Advantages: Offers predictability, regulatory flexibility, and a science-based foundation for process control. It enables proactive quality assurance and reduces batch failures by understanding and controlling variation sources.
- Applications: Ideal for well-characterized processes where mechanistic understanding is possible, such as the manufacturing of solid dosage forms and the application of PAT for real-time release [78].
Directed Evolution Approaches
Directed evolution, in contrast, mimics natural selection by employing iterative cycles of diversification and selection to arrive at optimized systems without requiring complete a priori knowledge of the underlying mechanisms [3]. In manufacturing, this parallels data-driven optimization strategies where machine learning algorithms iteratively refine process parameters based on performance outcomes.
- Key Methodology: The process involves two main steps: (1) Genetic Diversification: Introducing variation through methods like error-prone PCR or DNA shuffling in biological contexts, or through designed disturbance and parameter perturbation in process control; and (2) Isolation of Variants of Interest: Screening or selecting for improved performance using high-throughput methods like Fluorescence-Activated Cell Sorting (FACS) or MS-based methods in biology, or through real-time performance monitoring and model adaptation in automation [3].
- Advantages: Can optimize complex systems with non-linear interactions or incomplete mechanistic models. It is highly adaptable and can uncover novel solutions not predicted by existing models.
- Applications: Particularly valuable for optimizing complex bioprocesses with multiple interacting variables, such as upstream biomanufacturing using CHO cell cultures, and for handling raw material variability or process drift [3] [79].
Comparative Analysis of Advanced Control Technologies
This section objectively compares the performance of key technologies for advanced process control, focusing on their alignment with trait-based or directed evolution principles.
Process Analytical Technology (PAT) Tools
PAT tools are the sensory organs of a modern manufacturing plant, providing the real-time data essential for both control philosophies. The table below compares major PAT technologies used in solid dosage manufacturing [80] [78].
Table 1: Comparison of Process Analytical Technologies (PAT)
| Technology | Measured Attribute | Principle | In-line Applicability | Key Advantage |
|---|---|---|---|---|
| NIR Spectroscopy | Chemical composition, moisture content | Molecular overtone and combination vibrations | Excellent | Non-destructive, deep penetration |
| Raman Spectroscopy | Chemical structure, polymorphism | Inelastic scattering of monochromatic light | Good | Minimal sample preparation, specific |
| Spatial Filter Velocimetry | Particle size, velocity | Spatial filtering of scattered light | Excellent | Robust for particle size in flow |
| Ultrasonic Backscattering | Particle size, concentration | Scattering of high-frequency sound waves | Good | Effective in opaque systems |
| Terahertz Pulsed Imaging | Coating thickness, density | Time-domain spectroscopy | Limited | Deep penetration for coating analysis |
Model Predictive Control vs. Conventional PID Control
The core of advanced process control lies in the decision-making algorithm. The following table and experimental data compare the performance of MPC, a sophisticated trait-based method, against conventional Proportional-Integral-Derivative (PID) control.
Table 2: Performance Comparison of MPC vs. PID Control for a Bioreactor pH Control Loop [79]
| Performance Metric | PID Controller | MPC Controller | Improvement |
|---|---|---|---|
| Setpoint Tracking (7.10 to 7.15 pH) | ~35 minutes | ~15 minutes | 57% faster |
| Setpoint Tracking (7.10 to 7.07 pH) | ~22 minutes | ~4 minutes | 82% faster |
| Disturbance Rejection | Moderate | High | Superior constraint handling |
| Multivariable Capability | Limited (requires decoupling) | Native | Handious interactions directly |
| Computational Demand | Low | High | Requires powerful processor |
Experimental Protocol for Bioreactor Control [79]:
- System: A bioreactor for continuous biomanufacturing with CHO cell culture.
- Control Loops: pH, dissolved oxygen (DO), and temperature were controlled via inbuilt PI controllers, with a supervisory MPC for setpoint optimization.
- Monitoring: pH was monitored using an inbuilt electrode sensor. For MPC, a control-relevant process model was developed from experimental data.
- Implementation: The MPC system was implemented via a Distributed Control System (DCS) using a multi-layer communication protocol (OPC DA/UA) integrating DeltaV software with the bioreactor hardware.
- Performance Assessment: Controllers were evaluated for setpoint tracking by introducing step changes in pH and for disturbance rejection.
The data demonstrates MPC's significant advantage in dynamic response, a critical trait for maintaining processes within the design space and ensuring reproducibility despite disturbances.
Automation Platforms: MCP for Enterprise Integration
Automation platforms provide the nervous system that connects PAT sensors and MPC brains. The Model Context Protocol (MCP), an open standard developed by Anthropic, acts as a "USB-C port for AI applications," standardizing how AI models connect with tools and data sources [81] [82].
Table 3: Comparison of Enterprise MCP Automation Platforms
| Platform | Core Strength | Key Enterprise Feature | Performance | Integration Ecosystem |
|---|---|---|---|---|
| TrueFoundry | Purpose-built MCP orchestration | Centralized MCP Gateway with RBAC and audit logging | Sub-10ms latency, 350+ RPS | Pre-built servers for Slack, GitHub, Sentry |
| GitHub Copilot | Developer productivity | Native MCP for repository management & issue tracking | N/S | Tight integration with GitHub ecosystem |
| Microsoft Visual Studio | Deep IDE integration | Centralized configuration via group policies | N/S | Seamless with Azure DevOps, Teams, Office 365 |
| AWS SageMaker | Scalable ML infrastructure | Leverages mature AWS ML and data services | N/S | Deep integration with AWS service ecosystem |
N/S: Not Specified in the search results. The primary advantage of MCP is solving the "N×M" integration problem, where complexity grows exponentially with each new AI-system connection. By providing a standardized protocol, MCP platforms reduce development overhead and technical debt, enabling scalable and maintainable AI ecosystems [82]. This is foundational for implementing both trait-based control (by seamlessly integrating data flows from PAT to MPC) and directed evolution (by enabling AI agents to dynamically discover and use new data sources and tools).
The Scientist's Toolkit: Essential Research Reagent Solutions
The following table details key reagents, software, and hardware solutions essential for implementing advanced process control strategies.
Table 4: Essential Research Reagent Solutions for Advanced Process Control
| Item Name | Type | Function / Application | Relevant Context |
|---|---|---|---|
| KEGG Pathway Database | Software/Bioinformatics | A trait library for defining metabolic pathways and functional traits from genomic data [74]. | Trait-Based Approaches |
| TbasCO Software | Software/Bioinformatics | Identifies expression-based attributes of predefined traits using time-series transcriptomics data [74]. | Trait-Based Approaches |
| Error-Prone PCR Kit | Wet Lab Reagent | Introduces random mutations during gene amplification for library generation in directed evolution [3]. | Directed Evolution |
| Phylo-HMGP Model | Software/Bioinformatics | A continuous-trait probabilistic model for identifying genome-wide evolutionary patterns in functional genomic data [83]. | Directed Evolution |
| Raman Spectrometer | PAT Hardware | Provides molecular-level information for in-line monitoring of chemical attributes and real-time release [80] [78]. | PAT Implementation |
| DeltaV DCS | Automation Hardware | A distributed control system platform for implementing and orchestrating advanced control strategies like MPC [79]. | MPC & Automation |
| CHO Cell Line | Biological Reagent | Model system for monoclonal antibody production in upstream biomanufacturing process development [79]. | Biologics Manufacturing |
Workflow and Signaling Pathways
The implementation of an advanced control strategy is a multi-stage process. The diagram below outlines a generalized workflow integrating PAT, MPC, and MCP-based automation for a continuous manufacturing line.
Advanced Process Control Workflow
The workflow illustrates the continuous cycle of measurement, decision, and action. PAT sensors provide real-time data on CQAs and CPPs, which is processed and fed to the MPC. The MPC uses a process model to predict future states and compute optimal control decisions. These decisions are executed via the MCP automation platform, which orchestrates the process actuators. The MCP platform also enriches the context by dynamically discovering available data sources and tools, closing the loop for continuous quality assurance [82] [79] [78].
The pursuit of enhanced reproducibility in pharmaceutical manufacturing is best served by a strategic integration of PAT, MPC, and automation technologies. The comparative data presented in this guide clearly shows that MPC outperforms conventional PID control in dynamic response, leading to tighter process control. Furthermore, MCP-based automation platforms are critical for managing the complexity of integrated systems, reducing both development time and technical debt.
Framing this within the broader scientific context, trait-based approaches provide the necessary foundation for regulatory compliance and scientific understanding, making them the default for well-characterized unit operations and final product control. Meanwhile, directed evolution principles offer a powerful supplemental strategy for optimizing highly complex, non-linear, or poorly understood sub-processes, or for the continuous adaptation of models in response to process drift. The future of advanced process control lies not in choosing one philosophy over the other, but in leveraging their respective strengths—applying the rigorous, definitional power of trait-based control where possible, and harnessing the adaptive, exploratory power of directed evolution where necessary—within a seamlessly automated and data-rich manufacturing environment.
In both drug discovery and community optimization research, a fundamental challenge persists: how to design libraries that are both diverse enough to explore vast possibility spaces and small enough to be practically screenable. This challenge manifests differently across domains—from fragment-based drug discovery (FBDD) in pharmaceutical research to directed evolution in protein engineering—yet the core tension between diversity and practicality remains constant. In FBDD, libraries of small fragment-sized compounds (MW < 300) enable efficient exploration of chemical space, while in directed evolution, libraries of genetic variants probe sequence-function relationships [84] [3].
Across these fields, researchers must navigate the critical trade-off between library size and structural diversity. Larger libraries offer broader coverage but impose significant experimental burdens, whereas smaller, well-designed libraries can achieve comparable diversity with dramatically reduced screening requirements [84]. This article examines optimization strategies and quantitative frameworks that enable researchers to balance these competing demands effectively, comparing trait-based diversity approaches with directed evolution methodologies across multiple scientific domains.
Quantitative Metrics for Library Diversity and Size
Measuring Diversity in Fragment-Based Drug Discovery
In FBDD, diversity is quantified using structural descriptors that enable direct comparison of different library selections. Key metrics include:
- Tanimoto Similarity: Assesses pairwise structural similarity between compounds, with lower average similarity indicating greater diversity [84]
- Richness: The number of unique structural fingerprints present in a library, representing the absolute count of distinct structural features [84]
- True Diversity: A composite metric incorporating both the number of unique structural features and their proportional abundances, providing a more comprehensive diversity assessment [84]
Research demonstrates an interesting size-diversity relationship: while diversity generally increases with library size, there exists an optimal size point beyond which marginal diversity gains diminish or even become negative. Studies of commercially available fragments revealed that approximately 2,000 fragments (less than 1% of available compounds) can achieve the same true diversity level as all 227,787 available fragments, while maximum true diversity occurs at approximately 18,000 fragments (less than 8% of available compounds) [84].
Table 1: Size-Diversity Relationships in Fragment Libraries
| Library Size | Percentage of Total Fragments | Marginal Richness Efficiency | Achieved True Diversity |
|---|---|---|---|
| 100 | 0.04% | 28.9 fingerprints/compound | Low |
| 2,000 | 0.88% | Moderate | Equivalent to full library |
| 5,000 | 2.19% | 13.4 fingerprints/compound | High |
| 18,000 | 7.90% | Low/negative | Maximum |
| 227,787 | 100% | None | Reference point |
Diversity Metrics in Directed Evolution
In directed evolution, diversity assessment focuses on sequence space coverage and phenotypic variance. Unlike FBDD's structural fingerprints, directed evolution employs:
- Mutational Diversity: The range of amino acid substitutions at target positions
- Epistatic Interactions: Non-additive effects between mutations that create rugged fitness landscapes
- Functional Coverage: The proportion of functional variants within the library [3] [67]
The optimal library size in directed evolution depends on the complexity of the fitness landscape and the screening capacity. For epistatic landscapes (where mutation effects are non-additive), larger libraries are often necessary to capture beneficial combinations, though smart library design can reduce the required size [67].
Experimental Protocols for Library Design and Analysis
Protocol 1: Diversity-Optimized Fragment Library Design
Objective: Select a diverse subset of fragments from commercial collections that maximizes structural diversity while minimizing library size [84].
Materials:
- Database of commercially available fragments (e.g., ZINC database)
- Molecular fingerprinting software (e.g., extended-connectivity fingerprints)
- Diversity selection algorithm
- Clustering and partitioning tools
Methodology:
- Compound Retrieval: Retrieve structures of 227,787 commercially available fragments filtered by 'Rule-of-3' criteria (MW < 300, hydrogen bond donors ≤ 3, hydrogen bond acceptors ≤ 3, ClogP ≤ 3) [84]
- Descriptor Calculation: Generate structural fingerprints for all compounds using extended-connectivity fingerprints to represent molecular structures [84]
- Diversity Selection:
- Perform diversity-based selection using maximum dissimilarity algorithms
- Generate parallel random selections for comparison
- Create libraries with sizes ranging from 100 to 100,000 compounds
- Diversity Assessment:
- Calculate pairwise Tanimoto similarities
- Determine richness (number of unique fingerprints)
- Compute true diversity using Equation (1): D = 1/âˆ(pi^pi), where pi represents proportional abundance of the i-th fingerprint [84]
- Optimal Size Determination: Identify library size where marginal diversity gains diminish significantly
Validation: Compare hit rates from diversity-based versus random selections across multiple biological targets to confirm improved screening efficiency.
Protocol 2: Machine Learning-Assisted Directed Evolution
Objective: Optimize protein fitness through iterative rounds of mutagenesis and screening with machine learning guidance to reduce experimental burden [67].
Materials:
- Parent protein sequence
- Mutagenesis system (e.g., error-prone PCR, site-saturation mutagenesis)
- High-throughput screening assay
- Machine learning framework for protein fitness prediction
Methodology:
- Define Design Space: Select k target residues for optimization, creating a 20^k possible sequence space [67]
- Initial Library Construction: Generate initial variant library through saturation mutagenesis at all k positions using NNK degenerate codons [67]
- First-Round Screening:
- Express and purify variant proteins
- Measure fitness using target-specific assay (e.g., enzyme activity, binding affinity)
- Collect sequence-fitness data for 100-500 variants
- Machine Learning Model Training:
- Encode protein sequences using appropriate representations (one-hot, embedding, physicochemical properties)
- Train supervised model to predict fitness from sequence
- Implement uncertainty quantification (frequentist methods preferred over Bayesian for epistatic landscapes) [67]
- Iterative Optimization:
- Apply acquisition function to rank unscreened sequences
- Select top N variants (balancing exploration and exploitation)
- Screen selected variants experimentally
- Update model with new data
- Repeat for 3-5 rounds or until fitness convergence
Validation: Compare final variants with traditional directed evolution outcomes, assessing both fitness improvements and experimental resource requirements.
Trait-Based vs. Directed Evolution Approaches
The fundamental distinction between trait-based library design and directed evolution reflects different philosophical approaches to exploration and optimization.
Trait-Based Diversity Design
Trait-based approaches (including FBDD) prioritize systematic coverage of chemical or sequence space based on predefined structural or physicochemical descriptors [84] [85]. These methods:
- Rely on similar property principle: Structurally similar compounds likely have similar properties [85]
- Employ rational design using molecular descriptors including physicochemical properties, topological indices, and fingerprint-based descriptors [85]
- Utilize scaffold diversity analysis to ensure coverage of different chemotypes [85]
- Implement Pareto ranking for multi-objective optimization, balancing diversity with drug-like properties [85]
Directed Evolution Approaches
Directed evolution mimics natural evolutionary processes, emphasizing functional selection over predetermined diversity metrics [3] [47]. These methods:
- Prioritize functional outcomes over structural diversity
- Employ iterative exploration based on experimental feedback
- Leverage epistatic interactions that create cooperative effects [67]
- Utilize growth-coupled selection or screening-based methods depending on the system [47]
Table 2: Comparison of Library Design Approaches
| Parameter | Trait-Based Diversity Design | Directed Evolution |
|---|---|---|
| Primary Objective | Maximize structural/chemical diversity | Maximize functional fitness |
| Design Principle | Similar property principle | Evolutionary pressure |
| Diversity Metrics | Tanimoto similarity, richness, true diversity | Sequence coverage, functional variance |
| Optimization Approach | Rational design based on descriptors | Iterative experimental feedback |
| Library Size Strategy | Identify optimal diversity point | Match screening capacity |
| Epistasis Handling | Limited explicit consideration | Central to exploration strategy |
| Experimental Burden | Lower through rational design | Higher due to iterative screens |
| Success Measurement | Hit rates across multiple targets | Fitness improvement for specific goal |
Visualization of Library Design Workflows
Trait-Based Fragment Library Design
Trait-Based Library Design Workflow
Machine Learning-Assisted Directed Evolution
Directed Evolution with Machine Learning
The Scientist's Toolkit: Essential Research Reagents and Solutions
Table 3: Key Reagents for Fragment Library Design and Screening
| Resource | Function | Application Example |
|---|---|---|
| ZINC Database | Source of commercially available fragment structures | Retrieving 227,787 filtered fragments for library design [84] |
| Extended-Connectivity Fingerprints | Structural representation for diversity calculations | Generating molecular descriptors for similarity analysis [84] |
| Rule-of-3 Filters | Criteria for selecting fragment-like compounds | Pre-filtering compounds by MW < 300, HBD ≤ 3, HBA ≤ 3, ClogP ≤ 3 [84] |
| Groupwisdom Software | Concept mapping platform for stakeholder engagement | HPV vaccination strategy prioritization with community input [86] |
| Diversity Selection Algorithms | Computational methods for maximizing library diversity | Selecting optimal fragment subsets from large collections [84] |
Table 4: Key Reagents for Directed Evolution Campaigns
| Resource | Function | Application Example |
|---|---|---|
| Error-Prone PCR | Method for introducing random mutations across whole sequence | Creating diverse variant libraries in subtilisin E engineering [3] |
| NNK Degenerate Codons | Saturation mutagenesis approach covering all amino acids | ParPgb active site engineering at five residues [67] |
| Fluorescence-Activated Cell Sorting (FACS) | High-throughput screening based on fluorescence | Sortase engineering with product entrapment [3] |
| FoldX Suite | Protein design software for predicting thermodynamic stability | Calculating ΔΔG for structure-based regularization [87] |
| Active Learning-assisted DE (ALDE) | Machine learning framework for protein engineering | Optimizing ParPgb cyclopropanation yield from 12% to 93% [67] |
Discussion: Integration and Future Directions
The comparison between trait-based diversity design and directed evolution reveals complementary strengths that can be integrated into hybrid approaches. Trait-based methods excel in systematic exploration of chemical space, while directed evolution enables functional optimization in complex biological contexts.
Emerging strategies include:
- Structure-based regularization in machine learning-assisted directed evolution, which incorporates thermodynamic stability predictions to maintain protein fold integrity while optimizing function [87]
- Active learning frameworks that leverage uncertainty quantification to balance exploration of new sequence regions with exploitation of known high-fitness variants [67]
- Multi-objective optimization using Pareto ranking to simultaneously optimize diversity, drug-likeness, and other key properties [85]
These integrated approaches demonstrate that the dichotomy between trait-based and directed evolution strategies is increasingly blurring, with modern library design incorporating elements of both rational design and functional selection.
Optimizing library design requires careful balancing of diversity with practical screenable size. In fragment-based drug discovery, quantitative metrics reveal that 1-2% of available compounds can capture the majority of chemical diversity, dramatically reducing screening burdens. In directed evolution, machine learning assistance and smart library design strategies enable efficient navigation of complex fitness landscapes with minimal experimental iterations.
The choice between trait-based and directed evolution approaches depends on multiple factors: knowledge of the target space, availability of high-throughput assays, understanding of structure-function relationships, and resources for library construction and screening. By applying the quantitative frameworks, experimental protocols, and computational tools outlined in this comparison, researchers can make informed decisions to optimize their library design strategies across diverse applications in drug discovery and protein engineering.
Successful library design ultimately requires aligning methodology with project goals—whether prioritizing broad exploration of chemical space or focused optimization of specific functions—while leveraging appropriate metrics to balance diversity with practical screening constraints.
Strategies for Managing Unintended Fitness Costs and Off-Target Effects
In the pursuit of optimizing biological systems, researchers and drug developers face a fundamental challenge: how to achieve desired traits while minimizing unintended consequences. Two predominant philosophical approaches have emerged—trait-based optimization and directed evolution—each with distinct strengths, limitations, and risk profiles concerning unintended fitness costs and off-target effects. Trait-based optimization typically involves targeted genetic modifications aimed at specific phenotypic outcomes, often leveraging precise gene-editing tools like CRISPR/Cas9. In contrast, directed evolution harnesses principles of Darwinian selection in laboratory settings to iteratively improve biological functions through rounds of genetic diversification and screening [4] [88]. While both approaches have transformed biological engineering, their implementation carries different implications for the emergence and management of unintended effects that can compromise experimental outcomes and therapeutic applications.
Unintended fitness costs refer to reductions in organismal viability, reproductive success, or overall function resulting from genetic modifications, even when those modifications successfully confer the desired primary trait [89] [90]. Off-target effects encompass unintended genetic alterations at sites beyond the intended target, particularly relevant in CRISPR/Cas9 applications where non-specific DNA cleavage can disrupt functional genetic elements [91] [90]. Understanding and managing these unintended consequences is critical for developing effective therapeutics and engineered biological systems with predictable behavior and minimal adverse effects.
Analytical Framework: Comparing Optimization Approaches
Table 1: Comparison of Trait-Based versus Directed Evolution Approaches
| Parameter | Trait-Based Approaches | Directed Evolution Approaches |
|---|---|---|
| Core Principle | Rational design of specific genetic modifications | Iterative rounds of diversification and selection |
| Typical Tools | CRISPR/Cas9, homologous recombination | Error-prone PCR, DNA shuffling, gene shuffling |
| Fitness Cost Management | Post-hoc assessment; high-fidelity enzyme variants | Built-in through selective pressure during screening |
| Off-Target Effect Management | Computational guide RNA design; high-fidelity Cas9 variants | Functional screening inherently ignores silent off-target mutations |
| Key Advantages | Precision; speed for well-characterized targets | No requirement for structural knowledge; discovers cooperative mutations |
| Primary Limitations | Requires extensive target knowledge; prone to unanticipated fitness costs | Limited library diversity; screening throughput bottlenecks |
| Best Applications | Gene knockouts; specific point mutations; pathway disruption | Protein engineering; metabolic pathway optimization; novel function creation |
The selection between these approaches involves fundamental trade-offs. Trait-based methods offer greater precision but require comprehensive understanding of the system to avoid unintended consequences [91]. Directed evolution explores a broader mutational landscape and can identify non-intuitive solutions that simultaneously maintain fitness while achieving the desired function [4] [88]. For community optimization research in microbial systems or complex cellular environments, a hybrid approach often proves most effective, using directed evolution to identify beneficial mutations and trait-based methods to precisely incorporate them while monitoring for emergent fitness costs.
Experimental Evidence: Quantitative Assessment of Unintended Consequences
Fitness Costs in Trait-Based Approaches
CRISPR/Cas9 systems provide a powerful case study for examining unintended fitness costs in trait-based approaches. A comprehensive assessment in Drosophila melanogaster demonstrated that standard Cas9 expression imposed significant fitness costs primarily through off-target effects rather than direct costs of protein expression [90]. The study employed a sophisticated experimental design with four distinct constructs to disentangle different cost components:
- Cas9_gRNAs: Contained Cas9 and guide RNAs targeting a gene-free region
- Cas9_no-gRNAs: Contained Cas9 but no guide RNAs
- no-Cas9_no-gRNAs: Contained only fluorescent marker (control)
- Cas9HF1_gRNAs: Featured high-fidelity Cas9 variant with guide RNAs
Table 2: Quantified Fitness Costs of CRISPR/Cas9 Constructs in Drosophila
| Construct Type | Fitness Cost (Selection Coefficient) | Primary Cost Source | Competitive Outcome |
|---|---|---|---|
| Standard Cas9 with gRNAs | Significant (exact value inferred) | Off-target effects | Outcompeted by wild-type |
| Standard Cas9 without gRNAs | Minimal | Direct expression costs | Nearly equal to wild-type |
| Fluorescent marker only (Control) | None | N/A | Equal to wild-type |
| High-fidelity Cas9HF1 with gRNAs | Minimal | Largely eliminated | Similar to wild-type |
Researchers used a maximum likelihood framework to analyze allele frequency trajectories in cage populations, revealing that a model with no direct fitness costs but moderate costs due to off-target effects best fit the experimental data [90]. This finding was corroborated by individual fitness component assays measuring viability, fecundity, and mate choice. Importantly, the high-fidelity Cas9HF1 variant showed dramatically reduced fitness costs while maintaining efficient on-target activity, suggesting a viable strategy for mitigating unintended fitness consequences in trait-based approaches.
Fitness Costs in Antimicrobial Resistance
Studies of fungicide resistance in Colletotrichum siamense provide compelling evidence for context-dependent fitness costs associated with specific mutations. Using CRISPR/Cas9-mediated homology-directed repair, researchers introduced an E198A point mutation in β-tubulin that confers resistance to thiophanate-methyl in sensitive isolates [91]. The experimental protocol enabled precise comparison between genetically identical strains differing only at the target codon.
Of 41 comparisons across in vitro and detached fruit assays, mutant isolates appeared to be as fit as wild-type isolates in 24 comparisons (58.5%), and more fit in 10 comparisons (24.4%) [91]. This demonstrates that resistance mutations do not necessarily impose fitness costs and may even enhance fitness in certain environments, complicating resistance management strategies. The ribonucleoprotein (RNP) complex-mediated CRISPR/Cas9 system achieved an average transformation efficiency of 72% without detectable off-target mutations, highlighting the precision of modern trait-based approaches [91].
Unintended Consequences in Directed Evolution
Directed evolution campaigns have demonstrated remarkable success in improving protein properties while managing fitness costs through selective pressure. A landmark study evolving subtilisin E for enhanced activity in dimethylformamide achieved a 256-fold improvement after three rounds of error-prone PCR and screening [4] [88]. The resulting variant contained six cooperative mutations that collectively enhanced function without compromising stability.
The staggered extension process (StEP) for in vitro recombination has further demonstrated how directed evolution can optimize complex traits while maintaining fitness. Using this approach, researchers evolved subtilisin E to exhibit thermostability equal to its thermophilic homolog thermitase while maintaining enzymatic function [4]. This illustrates how directed evolution can identify mutational combinations that achieve desired traits while preserving overall protein fitness—a particular challenge for rational design approaches.
Methodological Guide: Experimental Strategies for Mitigation
Protocol for Assessing Fitness Costs in CRISPR/Cas9 Systems
The following experimental protocol, adapted from fitness cost assessments in Drosophila melanogaster [90], provides a robust framework for quantifying unintended consequences:
Construct Design: Develop multiple transgenic constructs to disentangle direct versus off-target fitness costs:
- Experimental construct (Cas9 + gRNAs)
- Expression control (Cas9 without gRNAs)
- Integration control (fluorescent marker only)
- High-fidelity variant (Cas9HF1 + gRNAs)
Transgenic Generation: Integrate constructs into identical genomic locations using site-specific recombination systems to control for position effects.
Competition Assays: House experimental and control genotypes together in replicated population cages under controlled conditions.
Frequency Monitoring: Track allele frequencies over 10+ generations using fluorescent markers or molecular genotyping.
Selection Coefficient Estimation: Apply maximum likelihood framework to allele frequency trajectories to quantify fitness costs.
Fitness Component Validation: Conduct individual assays for viability, fecundity, and mating success to verify population-level observations.
Off-Target Assessment: Whole-genome sequence representative samples to identify potential off-target mutations.
Experimental Workflow for Fitness Cost Assessment
Protocol for Directed Evolution with Fitness Constraints
This protocol, adapted from successful protein evolution campaigns [4] [88], incorporates fitness constraints during selection:
Library Generation:
- Apply error-prone PCR with tuned mutation rates (1-5 mutations/kb)
- Use DNA shuffling to recombine beneficial mutations
- Implement saturation mutagenesis at identified hotspots
Dual Selection Screening:
- Primary screen for desired functional improvement
- Counter-selection against fitness costs (e.g., growth rate, stability)
Iterative Enrichment:
- Isolate top performers from each round
- Use as templates for subsequent diversification
- Gradually increase selection stringency over rounds
Comprehensive Characterization:
- Measure kinetic parameters for desired function
- Assess fitness parameters under application conditions
- Determine structural impacts of beneficial mutations
Research Reagent Solutions
Table 3: Essential Research Reagents for Managing Unintended Effects
| Reagent/Category | Specific Examples | Function & Application |
|---|---|---|
| High-Fidelity Nucleases | Cas9HF1 [90], other engineered Cas variants | Reduces off-target effects in CRISPR-based approaches while maintaining on-target activity |
| Diversification Enzymes | Error-prone polymerases (Taq), DNaseI for shuffling [4] [88] | Creates genetic diversity for directed evolution campaigns |
| Selection Systems | Antibiotic resistance, fluorescence-activated cell sorting, auxotrophic markers [4] | Enables high-throughput screening of variant libraries |
| Assembly Reagents | Homology-directed repair templates, Gibson assembly mixes [91] | Facilitates precise genetic modifications and construct generation |
| Analytical Tools | Deep sequencing platforms, plate readers, flow cytometers [89] [90] | Quantifies intended and unintended effects of genetic modifications |
Integrated Management Strategy: A Hybrid Framework for Community Optimization
For community optimization research involving complex microbial consortia or cellular ecosystems, a hybrid approach that integrates both trait-based and directed evolution strategies provides the most robust framework for managing unintended fitness costs and off-target effects. This integrated methodology employs iterative design-build-test-learn (DBTL) cycles that leverage the strengths of both approaches while mitigating their respective limitations.
The proposed framework involves:
- Systematic Trait Identification: Use directed evolution to identify mutations that confer desired functions in model systems
- Precise Trait Integration: Employ trait-based methods to incorporate beneficial mutations into target community members
- Community-Level Fitness Assessment: Monitor emergent properties and stability in simplified synthetic communities
- Iterative Refinement: Apply lessons learned to inform subsequent engineering cycles
This approach is particularly valuable for optimizing microbial communities for therapeutic applications (e.g., live biotherapeutics) or industrial processes (e.g., consolidated bioprocessing), where both specific functions and overall community stability are critical success factors.
Hybrid Framework for Community Optimization
Effectively managing unintended fitness costs and off-target effects requires thoughtful selection and implementation of biological engineering strategies. Trait-based approaches benefit from precision but require vigilant monitoring of potential fitness costs and off-target effects through careful controls and high-fidelity reagents. Directed evolution offers powerful functional screening that inherently bypasses some unintended effects but faces limitations in library diversity and screening throughput. For the complex challenge of community optimization, a hybrid approach that leverages the complementary strengths of both methodologies provides the most promising path forward, enabling the development of robust, stable biological systems with minimized unintended consequences for therapeutic and industrial applications.
Head-to-Head Comparison and Validation of Engineered Systems
In the pursuit of optimizing biological systems for research and industrial applications, two dominant paradigms have emerged: trait-based approaches and directed evolution. Trait-based engineering relies on rational design, leveraging prior knowledge of biological components to assemble systems with desired functions. In contrast, directed evolution mimics natural selection in the laboratory, using iterative rounds of diversification and selection to steer biomolecules or organisms toward a predefined goal without requiring complete mechanistic understanding [3] [18]. This guide provides an objective comparison of these methodologies, focusing on key performance metrics, experimental protocols, and practical implementation resources to inform researchers in selection and application.
Core Principles and Conceptual Comparison
The foundational philosophies and operational mechanisms of trait-based and directed evolution approaches differ significantly, influencing their respective applications and outcomes.
Trait-Based Approaches are grounded in a rational, deductive framework. This methodology requires extensive prior knowledge of the system's components, such as the biochemical traits of amino acids in protein engineering or the metabolic capabilities of individual microbial strains in community assembly [9]. The engineer acts like a puzzle-solver, carefully selecting and combining these known pieces based on fundamental principles to achieve a target function. For instance, a synthetic microbial consortium might be constructed by combining species known to have complementary metabolic pathways, such as one species that hydrolyzes cellulose and another that ferments the resulting sugars into a desired product like bioethanol [9].
Directed Evolution, conversely, is an empirical, iterative process that harnesses the power of artificial selection. It does not require deep mechanistic knowledge of the system and instead remains agnostic to the underlying interactions between components (e.g., amino acids in a protein or species in a community) [3] [18] [9]. The process involves creating massive genetic diversity in a starting gene or population and then applying a high-throughput screening or selection pressure to isolate improved variants. These selected variants become the template for the next cycle of diversification and selection, leading to stepwise improvements [18]. A key advantage is its ability to discover non-obvious solutions that might be missed by rational design.
Table 1: Conceptual Comparison of the Two Approaches
| Feature | Trait-Based Approach | Directed Evolution |
|---|---|---|
| Underlying Philosophy | Rational, knowledge-based design | Empirical, blind-variation-and-selective-retention |
| Knowledge Requirement | High (e.g., structure, mechanism, traits) | Low to Moderate |
| Process Nature | Deductive, single-step assembly | Iterative (Diversification → Selection → Amplification) |
| Primary Driver | Researcher's hypothesis and design | Selective pressure and high-throughput screening |
| Typical Outcome Predictability | Theoretical, but often limited by complexity | Unpredictable, can lead to novel solutions |
Key Performance Metrics and Experimental Data
Evaluating the success of both approaches requires a multifaceted set of metrics. The choice of metrics is often dictated by the specific application, whether it be enzyme engineering, metabolic pathway optimization, or whole-cell biocatalyst development.
Quantitative Metrics for Assessment:
- Functional Activity/Productivity: This is the most direct metric, measuring the primary function of the engineered system. For enzymes, this could be catalytic efficiency (kcat/KM). For a microbial community, it is the titer, yield, and productivity of a target molecule (e.g., grams per liter per hour for 1,3-propanediol) [92].
- Stability: This encompasses robustness to environmental stresses such as high temperature, extreme pH, or the presence of organic solvents, which is critical for industrial processes [3] [18].
- Specificity: The ability to act on a target substrate while ignoring others is vital. This can be measured by substrate scope profiling or enantioselectivity (E value) in asymmetric synthesis [3].
- Binding Affinity: Particularly for binding proteins like antibodies, the dissociation constant (KD) is a key metric, often improved through affinity maturation [18].
- Fitness and Robustness: In community or whole-cell engineering, the growth rate, genetic stability, and resilience to evolutionary drift or invasion are crucial for sustained function [9].
Comparative Performance Insights: Computational frameworks like COSMOS have been developed to systematically compare the performance of different microbial systems. Such analyses reveal that the optimal choice between a simple monoculture (often a result of trait-based rational design) and a more complex co-culture (which can be optimized via directed evolution) is highly context-dependent. Key findings include [92]:
- Environmental Dependency: Microbial co-cultures often achieve their highest productivity advantage over the best-performing monoculture in anaerobic, nutrient-rich environments. This is attributed to enhanced metabolite exchange and more balanced growth rates between community members under these conditions.
- Product Specificity: The performance is also product-dependent. For instance, a co-culture of Shewanella oneidensis and Klebsiella pneumoniae was computationally predicted and experimentally validated as the most efficient system for producing 1,3-propanediol under anaerobic conditions [92].
- Functional Diversity Trade-off: On evolutionary timescales, a higher number of species (species diversity) does not automatically lead to greater functional diversity. Theory and data from land snail communities show that in tightly packed, species-rich communities, individual species evolve narrower trait breadths to avoid competition, which can paradoxically reduce the community's overall functional diversity [2].
Table 2: Key Quantitative Metrics for Evaluation
| Metric Category | Specific Measurable | Typical Experimental Method | Application Example |
|---|---|---|---|
| Activity & Productivity | Catalytic Efficiency (kcat/KM), Product Titer (g/L), Yield (g/g substrate) | Enzyme kinetics, HPLC/GC analysis | Comparing substrate-specific enzyme variants [3] |
| Stability | Melting Temperature (Tm), Half-life at operational condition | Thermofluor assay, circular dichroism | Selecting thermostable lipases for industrial processes [3] [18] |
| Specificity | Enantiomeric Excess (e.e.), Substrate Scope Profile | Chiral HPLC, mass spectrometry | Evolving transaminases for chiral amine synthesis [3] |
| Binding Affinity | Dissociation Constant (KD) | Surface Plasmon Resonance (SPR), Bio-Layer Interferometry (BLI) | Affinity maturation of therapeutic antibodies [18] |
| System Performance | Relative Productivity (vs. top monoculture) | Computational modeling (e.g., COSMOS), bioreactor runs | Identifying optimal microbial system for a product [92] |
Detailed Experimental Protocols
The implementation of trait-based and directed evolution approaches involves distinct, well-established workflows. Below are generalized protocols for each.
Trait-Based Microbial Consortium Assembly
This protocol outlines the rational design and construction of a synthetic microbial community [9].
Trait Identification and Selection:
- Objective: Define the target function (e.g., production of compound X from substrate Y).
- Procedure: Mine literature and databases for microbial species with known, complementary traits relevant to the objective. For example, select Species A for its ability to degrade a complex polymer and Species B for its capacity to convert the resulting monomers into the target product.
- Validation: Confirm traits in-house via growth assays and metabolite analysis.
Consortium Design and Modeling:
- Procedure: Use metabolic modeling (e.g., constraint-based reconstruction and analysis) to predict interactions and potential bottlenecks.
- Output: A predicted optimal ratio of starting species and environmental conditions (e.g., medium composition).
Experimental Assembly and Testing:
- Procedure: Cultivate selected species individually and then combine them in co-culture at the predicted ratio.
- Control: Run parallel monocultures of each species.
- Analysis: Monitor community composition (e.g., via plating, qPCR, or flow cytometry) and measure target function productivity over time.
General Directed Evolution Workflow
This protocol describes the iterative cycle for evolving improved biomolecules, such as enzymes [3] [18] [93].
Library Generation (Diversification):
- Objective: Create a large library of genetic variants of the starting gene.
- Common Methods:
- Error-Prone PCR: Uses reaction conditions that reduce the fidelity of DNA polymerase to introduce random point mutations across the entire gene [3].
- DNA Shuffling: Fragments and recombines homologous genes from different parents to create chimeric variants, useful for exploring recombined sequence space [3] [18].
- Saturation Mutagenesis: Targets specific residues (e.g., in an enzyme's active site) to explore all possible amino acid substitutions at those positions, creating a "focused library" [3].
Screening or Selection (Selection):
- Objective: Identify the rare, improved variants from the large library.
- Screening: Each variant is individually expressed and assayed, often using colorimetric or fluorogenic substrates in a high-throughput microtiter plate format. This provides quantitative data but has lower throughput [3] [18].
- Selection: Couples the desired function (e.g., enzyme activity) to cell survival or antibiotic resistance. This allows for evaluating extremely large libraries (millions to billions of variants) but is more complex to engineer and provides less quantitative information on the variants' distribution [18].
Gene Amplification:
- Objective: Isolate the genes of the best-performing variants to serve as templates for the next round.
- Procedure: For screening, plasmids are isolated from selected clones. For in vitro methods, the genes of functional variants are recovered via PCR.
- Next Step: The amplified genes are used to start the next round of diversification and selection, creating an iterative optimization cycle.
The following diagram visualizes the core, iterative process of a directed evolution experiment.
The Scientist's Toolkit: Essential Research Reagents and Materials
Successful implementation of these engineering strategies relies on a suite of specialized reagents and tools.
Table 3: Essential Research Reagents and Solutions
| Reagent / Material | Function / Description | Primary Application |
|---|---|---|
| Error-Prone PCR Kit | A ready-to-use mixture for performing PCR with low-fidelity polymerases to introduce random mutations. | Directed Evolution: Library Generation [3] |
| Phage/Yeast Display System | A platform where proteins are displayed on the surface of phages or yeast cells, allowing binding-based selection from vast libraries. | Directed Evolution: Selection of binders (e.g., antibodies) [3] [18] |
| Fluorescent/Achromogenic Substrate | A substrate that produces a detectable signal (color/fluorescence) upon enzyme action, enabling high-throughput screening. | Directed Evolution: Screening enzymatic activity [3] |
| Specialized Microbial Growth Media | Chemically defined or rich media tailored to support specific microbial functions or co-culture stability. | Trait-Based & Directed Evolution: Cultivation and selection [92] [94] |
| Metabolic Model (e.g., Genome-Scale Model) | A computational reconstruction of an organism's metabolism used to predict growth, product yield, and interactions. | Trait-Based Approach: Rational consortium design [92] [9] |
Visualization of Strategic Selection
The decision to use a trait-based approach or directed evolution is not mutually exclusive and can be guided by a logical assessment of the research context. The following diagram outlines key decision points.
Integrated and Future Perspectives
The distinction between trait-based and directed evolution is increasingly blurred by integrated "semi-rational" strategies. Focused libraries, which use structural knowledge to restrict randomization to key regions, combine the targeted efficiency of rational design with the exploratory power of evolution [3] [18]. Furthermore, computational tools are playing an ever-larger role. Environmentally focused strategies rationally manipulate factors like temperature and pH to optimize microbial function, a top-down approach applicable to both natural and engineered systems [94]. Advanced frameworks like COSMOS leverage dynamic modeling to simulate and predict the performance of monocultures versus co-cultures under specified conditions, providing a data-driven starting point for experimental efforts [92].
The future of biological optimization lies in the intelligent integration of these approaches. Leveraging computational predictions to inform rational design, and using directed evolution to refine and optimize these designs, creates a powerful, iterative engineering loop. This synergistic framework will accelerate the development of robust biocatalysts and microbial consortia for advanced therapeutic and biomanufacturing applications.
The optimization of biological communities, whether for therapeutic development or ecosystem engineering, hinges on a fundamental dichotomy in research approaches: trait-based strategies versus directed evolution. Trait-based approaches operate on the principle that detailed knowledge of functional traits—morphological, physiological, or ecological characteristics influencing organismal fitness and ecosystem functioning—enables predictive design and manipulation of communities [95] [96]. This methodology seeks to understand and harness the "value and the range of those species and organismal traits that influence ecosystem functioning" to achieve desired outcomes [96]. In contrast, directed evolution mimics natural selection in controlled settings, applying selective pressures to evolve populations toward optimized performance without requiring prior mechanistic knowledge of underlying traits [97]. Within this conceptual framework, functional diversity emerges as a critical bridging concept, representing the variety of organismal traits within a community that directly influence ecosystem dynamics, stability, productivity, and other aspects of ecosystem functioning [96].
This guide provides a comparative analysis of these competing paradigms through the lens of functional diversity validation. We objectively evaluate their performance across multiple research contexts, supported by experimental data and detailed methodologies, to inform strategy selection by researchers and drug development professionals engaged in community optimization research.
Theoretical Foundations: Defining and Quantifying Functional Diversity
Functional diversity is quantitatively distinct from, though related to, species richness. It measures the range and distribution of functionally relevant traits within a community, which directly impact ecosystem processes [96]. A community with high functional diversity typically exhibits a greater variety of resource use strategies, potentially leading to more stable and productive ecosystems [96].
The relationship between species diversity and functional diversity is not always positive or straightforward. Eco-evolutionary models demonstrate that in tightly-packed, species-rich communities, competition can force species to evolve narrower trait breadths to minimize overlap with neighbors [98]. This process can result in a negative relationship between species diversity and functional diversity, challenging the intuitive assumption that more species automatically guarantee greater functional variety [98].
Key Metrics and Measurement Approaches
Table 1: Functional Diversity Metrics and Their Applications
| Metric Name | Measurement Focus | Research Context | Interpretation |
|---|---|---|---|
| Rao's Q | Trait dissimilarity within a community | Remote sensing of global biomes [70] | Higher values indicate greater functional diversity; shows lower seasonal variation |
| Functional Richness | Range of trait values in a community | Global biome comparison [70] | Higher values indicate broader trait ranges; exhibits strong seasonal variation |
| Functional Evenness | Regularity of trait distribution | Ecosystem functioning studies [96] | Even distributions suggest optimal resource use |
| Functional Divergence | Degree of abundance in extreme traits | Community assembly studies [96] | High values indicate specialization in unusual traits |
Measurement strategies vary significantly between approaches. Trait-based methods often employ detailed characterization of specific functional traits, while directed evolution approaches may use high-throughput screening of bulk community properties. Remote sensing technologies now enable mapping of functional diversity patterns across large spatial and temporal scales, revealing pronounced seasonal dynamics across major biomes [70]. These temporal patterns highlight that functional diversity is not static but responds to environmental cycles and phenological changes, necessitating multi-temporal assessment for accurate characterization [70].
Comparative Analysis: Trait-Based Versus Directed Evolution Approaches
Performance Comparison Across Research Contexts
Table 2: Approach Comparison in Community Optimization Research
| Research Parameter | Trait-Based Approach | Directed Evolution Approach |
|---|---|---|
| Predictive Capability | High when trait-function relationships are well-established [96] | Limited a priori prediction, emerges from selection process [97] |
| Experimental Timescale | Often shorter once functional traits are identified [95] | Typically longer due to multiple evolutionary cycles [97] |
| Novelty Generation | Limited to natural trait variation or designed extensions [97] | High potential for novel combinations and emergent properties [99] |
| Measurement Requirements | Requires detailed trait characterization [95] [96] | Often relies on high-throughput screening [97] |
| Dependence on Prior Knowledge | High dependency on established trait databases [95] | Lower dependency, can discover unknown relationships [99] [97] |
| Success with Complex Traits | Effective for simple, well-conserved traits [95] | More effective for complex, polygenic traits [97] |
Empirical Validation Studies
Plant Defense Gene Analysis (Trait-Based)
A comprehensive analysis of nucleotide-binding site (NBS) domain genes across 34 plant species identified 12,820 NBS-domain-containing genes classified into 168 architectural classes, revealing both classical and species-specific structural patterns [100]. This trait-based study employed expression profiling to demonstrate upregulation of specific orthogroups (OG2, OG6, OG15) in different tissues under biotic and abiotic stresses [100].
Key Experimental Protocol:
- Gene Identification: Used PfamScan.pl HMM search script with default e-value (1.1e-50) against Pfam-A_hmm model to identify NBS domains [100]
- Classification System: Applied domain architecture classification following established methods, grouping genes with similar domain architectures into classes [100]
- Evolutionary Analysis: Conducted orthogroup analysis using OrthoFinder v2.5.1 with DIAMOND for sequence similarity and MCL for clustering [100]
- Expression Validation: Utilized RNA-seq data from IPF database and NCBI BioProjects, processing data through transcriptomic pipelines to generate expression heat maps [100]
- Functional Validation: Implemented virus-induced gene silencing (VIGS) of GaNBS (OG2) in resistant cotton to demonstrate its role in virus tolerance [100]
The research identified significant genetic variation between susceptible (Coker 312) and tolerant (Mac7) cotton accessions, with 6,583 unique variants in Mac7 and 5,173 in Coker312 NBS genes [100]. Protein-ligand interaction studies revealed strong binding of putative NBS proteins with ADP/ATP and core proteins of the cotton leaf curl disease virus [100].
AI-Driven Protein Design (Directed Evolution)
Cutting-edge research demonstrates how artificial intelligence models can now generate functional de novo proteins through semantic design strategies. The Evo genomic language model learns distributional semantics across prokaryotic genes to perform function-guided design of novel sequences [99]. This approach represents an advanced form of directed evolution, leveraging AI to accelerate the exploration of sequence-function space beyond natural evolutionary constraints [99].
Key Experimental Protocol:
- Model Architecture: Evo 1.5 model pretrained on 450 billion tokens from prokaryotic DNA sequences in OpenGenome, processing long genomic sequences at single-nucleotide resolution [99]
- Prompt Engineering: Curated eight prompt types including toxin/antitoxin sequences, their reverse complements, and upstream/downstream genomic contexts [99]
- Sequence Generation: Performed sampling using prompts, followed by filtering for sequences encoding protein pairs with in silico predicted complex formation [99]
- Novelty Filtering: Applied novelty filters requiring limited sequence identity to known proteins [99]
- Functional Validation: Implemented growth inhibition assays to quantify toxin activity, demonstrating approximately 70% reduction in relative survival for novel toxin EvoRelE1 [99]
This AI-augmented directed evolution approach successfully generated functional toxin-antitoxin pairs and anti-CRISPR proteins, including de novo genes with no significant sequence similarity to natural proteins [99]. The model demonstrated robust predictive performance, achieving over 80% protein sequence recovery for target genes based solely on operonic neighbors [99].
Research Strategy Workflow: Comparing fundamental approaches to community optimization.
The Scientist's Toolkit: Essential Research Reagents and Solutions
Table 3: Key Research Reagents for Functional Diversity Studies
| Reagent/Solution | Primary Function | Research Context |
|---|---|---|
| PfamScan HMM Script | Identification of protein domains from sequence data | Plant NBS gene discovery [100] |
| OrthoFinder Pipeline | Orthogroup inference and comparative genomics | Evolutionary analysis of gene families [100] |
| RNA-seq Databases (IPF) | Gene expression profiling under various conditions | Differential expression analysis [100] |
| VIGS Constructs | Virus-induced gene silencing for functional validation | Plant gene functional characterization [100] |
| Evo Genomic Model | AI-driven generation of novel functional sequences | De novo protein design [99] |
| Growth Inhibition Assays | Quantification of toxin activity in cellular systems | Validation of toxin-antitoxin systems [99] |
| Hyperspectral Imaging Data | Remote assessment of plant functional traits | Global functional diversity monitoring [70] |
| EnMAP Satellite Data | Spaceborne hyperspectral imagery | Multi-seasonal functional diversity mapping [70] |
Integration Strategies and Future Directions
The research paradigm is increasingly shifting toward integrated approaches that combine trait-based knowledge with directed evolution principles. AI-driven models now leverage genomic context to perform "semantic design" of novel functional sequences, effectively bridging the gap between mechanistic understanding and evolutionary exploration [99]. These integrated frameworks demonstrate robust success rates even for sequences with no significant similarity to natural proteins, highlighting the potential for accessing entirely novel regions of functional space [99] [97].
For research requiring rapid optimization of complex communities with limited prior knowledge, directed evolution approaches provide powerful discovery tools. Conversely, when detailed mechanistic understanding is available or required for regulatory approval, trait-based methods offer superior predictability and control. The emerging generation of AI-augmented tools promises to further blur these distinctions, creating new opportunities for community optimization across therapeutic development, biotechnology, and ecosystem management.
Strategy Selection Guide: Decision framework for selecting optimal research approaches based on available knowledge and data resources.
The engineering of biological systems for research and therapeutic purposes predominantly leverages two powerful paradigms: trait-based engineering and directed evolution. The former, often called rational design, involves the precise, knowledge-driven modification of an organism's genetic blueprint to instill a predefined trait or function. The latter, Synthetic Directed Evolution (SDE), mimics natural evolution in the laboratory through iterative cycles of diversification and selection to arrive at optimized biological molecules. Framed within a broader thesis on community optimization research, this guide provides an objective, data-driven comparison of these approaches, delineating their respective strengths, limitations, and ideal applications for researchers and drug development professionals.
Methodology at a Glance: Core Principles and Workflows
Trait-Based Engineering
Trait-based engineering is a targeted approach that relies on existing knowledge of gene function and regulatory mechanisms. The core principle is to directly introduce or modify specific genetic sequences to achieve a predetermined outcome. This approach has been powerfully enabled by CRISPR-Cas9 systems, which function as programmable "molecular scissors" to create double-strand breaks in DNA at precise locations, leading to targeted genetic modifications through cellular repair pathways like Non-Homologous End Joining (NHEJ) or Homology-Directed Repair (HDR) [101] [102]. More recently, this field has been revolutionized by the integration of Artificial Intelligence (AI), which uses large-language models trained on vast biological datasets to design novel, highly functional genome editors and biological parts from scratch, bypassing evolutionary constraints [103] [104].
The following diagram illustrates a generalized workflow for the AI-driven trait-based engineering approach:
Synthetic Directed Evolution (SDE)
SDE is an iterative, empirical approach that does not require prior mechanistic knowledge of the system. Its foundational principle involves creating a diverse library of genetic variants and applying a selective pressure to isolate individuals with improved or novel functions [15]. This process typically involves two main stages: a diversification stage, where mutations are introduced into target gene sequences (via methods like error-prone PCR, DNA shuffling, or CRISPR-based mutagenesis), and a selection stage, where the host population is screened or selected under specific conditions to identify improved variants [15]. Successful variants undergo multiple rounds of this cycle to accumulate beneficial mutations.
The diagram below outlines the core, iterative cycle of a Synthetic Directed Evolution experiment:
Comparative Performance Analysis
The following tables provide a side-by-side comparison of the two approaches across key performance and application metrics.
Table 1: Comparison of Methodological Strengths and Limitations
| Aspect | Trait-Based Engineering | Synthetic Directed Evolution (SDE) |
|---|---|---|
| Core Principle | Rational, knowledge-driven design of specific genetic changes [103] | Empirical, iterative cycles of diversification and selection [15] |
| Knowledge Requirement | High (requires understanding of gene function & regulatory mechanisms) [103] | Low (does not require prior knowledge of gene structure) [15] |
| Precision & Control | High; enables precise modifications with unmatched accuracy [101] [104] | Low; introduces random mutations, control is exerted via selection, not design |
| Development Speed | Potentially fast for well-understood traits; AI can accelerate design [104] | Can be slower due to multiple necessary rounds of iteration [15] |
| Exploration of Sequence Space | Limited to known or rationally designed variations | Vast; capable of discovering novel, non-obvious solutions [15] |
| Primary Risk | Design failures due to incomplete biological knowledge | Resource-intensive; potential for false positives during screening [15] |
Table 2: Experimental and Application-Based Comparison
| Aspect | Trait-Based Engineering | Synthetic Directed Evolution (SDE) |
|---|---|---|
| Key Tools/Technologies | CRISPR-Cas9, Base Editors, AI-generated editors (OpenCRISPR-1) [101] [104] | Error-prone PCR, DNA shuffling, MAGE, CRISPR-directed evolution [15] |
| Ideal for Unknown Targets | Poor (relies on known targets) | Excellent (can probe function without mechanistic insight) [15] |
| Typical Applications | Gene therapy (correcting pathogenic mutations), creating specific animal models, crop trait engineering [101] [102] | Enzyme engineering, optimizing metabolic pathways, improving protein stability, herbicide resistance in crops [15] |
| Throughput & Resource Demand | Lower throughput per experiment, but more targeted | High-throughput screening is often required, demanding significant resources [15] |
| Integration with AI/ML | AI used for de novo design of editors and components [104] | ML used to predict best-performing variants from sequence-activity data, guiding library design [15] |
Experimental Protocols in Practice
Protocol for AI-Driven Trait-Based Engineering
The following protocol outlines the steps for designing and deploying an AI-generated genome editor, as demonstrated for OpenCRISPR-1 [104].
- Data Curation and Model Training: Systematically mine terabases of genomic and metagenomic data to create a curated dataset of CRISPR operons (e.g., the CRISPR–Cas Atlas). Fine-tune a large language model (e.g., ProGen2) on this dataset to learn the sequence-function relationships of CRISPR-Cas proteins [104].
- AI-Based Protein Generation: Use the fine-tuned model to generate millions of novel Cas protein sequences. Filter these sequences for viability and cluster them to ensure diversity. The generated sequences are typically 40-60% identical to any known natural protein [104].
- In Silico Validation: Predict the 3D structures of top candidate proteins using tools like AlphaFold2 to confirm they adopt correct functional folds [104].
- Delivery and Testing in Human Cells:
- Delivery: Package the candidate editor's coding sequence and its guide RNA (sgRNA) into a suitable delivery vector (e.g., AAV, lentivirus, or lipid nanoparticles) and transfect into human cell lines [102] [104].
- Evaluation: Measure editing efficiency at the target locus using next-generation sequencing. Assess specificity by sequencing potential off-target sites to ensure the editor does not cleave unintended genomic regions [104].
Protocol for CRISPR-Mediated Directed Evolution (e.g., CasPER)
This protocol details the Cas9-mediated protein evolution reaction (CasPER), a specific SDE method for evolving enzymes in their native genomic context [15].
- Library Construction: Use error-prone PCR to introduce random mutations into a 300-600 bp target region of the gene of interest, generating a combinatorial library of mutagenized linear DNA donor fragments [15].
- Genomic Integration: Co-transform the library of mutagenized DNA donors into a host cell (e.g., yeast or bacteria) along with a plasmid expressing CRISPR-Cas9. The Cas9 enzyme induces a double-strand break at the precise genomic target, stimulating the host's homology-directed repair machinery to integrate the mutagenized donor fragments [15].
- Selection and Screening: Grow the population of engineered cells under a specific selective pressure (e.g., the presence of an herbicide for evolving tolerance, or a substrate for improving an enzyme's activity). Only variants with enhanced function will survive or grow preferentially [15].
- Variant Recovery and Iteration: Isolate the genetic material from the selected population. The selected variants can be used as a template for further rounds of diversification and selection to accumulate additional beneficial mutations [15].
The Scientist's Toolkit: Essential Research Reagents
Table 3: Key Reagents and Resources for Trait-Based and Directed Evolution Approaches
| Reagent/Resource | Function | Primary Application |
|---|---|---|
| CRISPR-Cas9 System | Programmable RNA-protein complex for creating targeted double-strand breaks in DNA [101] [102] | Foundational tool for both approaches (for precise editing or library generation) |
| AI-Designed Editor (e.g., OpenCRISPR-1) | A novel, highly functional Cas protein generated by a language model, offering potential improvements in activity/specificity [104] | Trait-Based Engineering |
| Error-Prone PCR Kit | Reagents for PCR that introduce random mutations during amplification, creating diverse variant libraries [15] | Synthetic Directed Evolution |
| Viral Delivery Vectors (AAV, LV) | Engineered viruses used to efficiently deliver CRISPR components into target cells, especially for in vivo applications [102] | Trait-Based Engineering |
| Non-Viral Delivery Vectors (LNPs) | Lipid nanoparticles that encapsulate and deliver CRISPR ribonucleoproteins or mRNA, avoiding immunogenicity concerns of viral vectors [102] | Trait-Based Engineering |
| Selection Agents (e.g., Herbicides, Antibiotics) | Chemical compounds applied to exert selective pressure, enriching for cellular populations with desired traits like resistance [15] | Synthetic Directed Evolution |
| CRISPR–Cas Atlas | A comprehensive, curated database of CRISPR operons used for training AI models to design new editors [104] | Trait-Based Engineering (AI-driven) |
The choice between trait-based engineering and synthetic directed evolution is not a matter of which is universally superior, but which is optimal for a specific research goal within a community optimization framework. Trait-based engineering excels when the biological mechanism is well-understood, the target is known, and the goal is precise, predictable modification. The advent of AI-driven design has dramatically expanded its potential, enabling the creation of novel biological tools that transcend natural diversity. Conversely, synthetic directed evolution is the method of choice for exploring the unknown, optimizing complex phenotypes, or discovering novel solutions when mechanistic insight is lacking. Its power lies in its ability to empirically test a vast landscape of possibilities. For the modern researcher, the most powerful strategy may often be a hybrid one, leveraging the exploratory power of SDE to identify promising variants and the precision of trait-based engineering to refine and implement them, all while utilizing AI and robust benchmarking to accelerate the entire cycle of discovery and application.
In the quest to optimize biological systems—from single enzymes to microbial consortia—researchers primarily navigate two powerful paradigms: trait-based approaches and directed evolution (DE). Trait-based strategies operate on a rational design principle, leveraging known functional traits, structural information, or phylogenetic data to predict and construct optimal biological configurations [9] [76]. In contrast, directed evolution mimics natural selection in a laboratory setting, using iterative rounds of random mutagenesis and high-throughput screening to empirically discover enhanced functions without requiring prior mechanistic knowledge [3] [88]. While often perceived as distinct, the integration of these methodologies is forging a robust, hybrid framework for community optimization research. This guide provides a comparative analysis of these strategies, detailing their experimental protocols, performance data, and practical implementation for scientific and drug development applications.
Methodological Comparison: Principles, Advantages, and Limitations
The following table provides a systematic comparison of the core methodologies, highlighting their strategic advantages and inherent constraints.
Table 1: Comparative Analysis of Trait-Based and Directed Evolution Approaches
| Aspect | Trait-Based Approaches | Directed Evolution |
|---|---|---|
| Core Principle | Rational design based on known traits, structure, or phylogeny [9] [76] | Laboratory mimicry of natural evolution through iterative diversification and selection [3] [88] |
| Required Prior Knowledge | High (e.g., structural data, ecological traits, mechanistic insights) [76] | Low to none; can proceed agnostically [105] [88] |
| Typical Workflow | Hypothesis-driven, bottom-up assembly or top-down manipulation [9] | Empirical, cyclic process of mutagenesis and screening/selection [3] |
| Key Advantage | Predictable, targeted interventions; deeper mechanistic understanding [76] | Discovers non-intuitive and highly effective solutions beyond rational design [88] |
| Primary Limitation | Limited by depth of functional understanding; can miss emergent properties [9] | High-throughput screening is a major bottleneck; can be labor-intensive [3] [105] |
| Best Suited For | Optimizing systems with well-characterized components and interactions [76] | Optimizing complex traits with unknown genetic basis or for generating novel functions [88] |
Experimental Protocols and Workflows
Trait-Based Community Assembly and Optimization
Trait-based approaches in microbial ecology involve constructing synthetic consortia from individual species with known metabolic or physiological traits [9] [76].
- Trait Identification and Characterization: Culture individual microbial strains in isolation. Quantify key functional traits such as substrate utilization profiles (e.g., using Biolog EcoPlates), growth rates under different conditions, metabolic byproduct secretion, and resistance to environmental stressors [76].
- In Silico Modeling and Prediction: Use the collected trait data to construct metabolic models or interaction networks. The goal is to predict combinations of strains that will exhibit desired community-level functions, such as efficient division of labor, cross-feeding (syntrophy), or enhanced ecosystem stability [9].
- Consortium Assembly: Co-culture the selected strains in a defined medium. The initial ratios can be informed by the predictive models.
- Function Monitoring and Refinement: Monitor the target community function over time (e.g., product yield, substrate degradation rate, community stability). The composition can be adjusted by changing media conditions or re-inoculation to steer the community toward the optimal state [9].
Core Directed Evolution Workflow for Protein Engineering
The standard directed evolution pipeline is an iterative cycle of diversity generation and screening [3] [88].
- Library Generation through Mutagenesis:
- Error-Prone PCR (epPCR): A standard method for random mutagenesis. The PCR reaction is made error-prone by using a non-proofreading polymerase (e.g., Taq polymerase), imbalanced dNTP concentrations, and the addition of manganese ions (Mn²âº) to achieve a target mutation rate of 1-5 mutations per kilobase [106] [88].
- DNA Shuffling: A recombination-based method. DNaseI randomly fragments a pool of related parent genes. The fragments are then reassembled in a primer-free PCR, allowing crossovers that shuffle mutations from different parents into novel combinations [88].
- Site-Saturation Mutagenesis: A semi-rational technique targeting specific amino acid positions. A specific codon is replaced with a degenerate codon (e.g., NNK, where N is any base and K is G or T) to generate all 19 possible amino acid substitutions at that site [3] [88].
- Screening and Selection:
- Microtiter Plate Screening: Individual library variants are expressed in a host (e.g., E. coli) and cultured in 96- or 384-well plates. Enzyme activity is assayed using colorimetric or fluorometric substrates, with a plate reader quantifying the signal [3] [88]. This is medium-throughput but provides quantitative data.
- Fluorescence-Activated Cell Sorting (FACS): An ultra-high-throughput method. Used when enzyme activity can be linked to the generation or loss of fluorescence, allowing for the screening of millions of cells in hours [3].
The following diagram illustrates the core, iterative cycle of a directed evolution experiment.
Diagram Title: The Core Directed Evolution Cycle
Performance Data and Comparative Outcomes
The effectiveness of each strategy is best illustrated by their application to real-world optimization challenges. The table below summarizes experimental data and outcomes from key application areas.
Table 2: Experimental Outcomes of Trait-Based and Directed Evolution Strategies
| Application Area | Strategy Employed | Experimental Outcome / Performance Gain | Key Methodology Details |
|---|---|---|---|
| Biofuel Synthesis | Directed Evolution [105] | Improvement of hydrocarbon-producing enzymes (e.g., cytochrome P450 OleTJE) for higher alkene/alkane yields. | Error-prone PCR and high-throughput screening using product-specific assays or biosensors [105]. |
| Industrial Biocatalysis | Directed Evolution [88] | Generation of subtilisin E variants with enhanced stability in harsh detergents. | Colony screening on milk-agar plates; active variants formed clear hydrolysis halos [88]. |
| Synthetic Microbial Consortia | Trait-Based [9] | Two-species system for bioethanol production from cellulose. | Leveraged native traits of C. phytofermentans (cellulose hydrolysis) and E. coli (fermentation) in a co-culture [9]. |
| Therapeutic Protein Engineering | Directed Evolution [88] | Development of therapeutic antibodies and viral vectors with improved binding affinity or specificity. | Phage display or yeast display (selection-based techniques) screening libraries >109 in size [88]. |
| Community Function Optimization | Trait-Based [76] | Enhanced ecosystem functions like nitrogen cycling or organic matter decomposition. | Community trait mean (weighted by species abundance) correlated with and predictive of process rates [76]. |
The Scientist's Toolkit: Essential Research Reagents and Solutions
Successful implementation of these strategies relies on a suite of specialized reagents and tools.
Table 3: Key Research Reagent Solutions for Trait-Based and Directed Evolution Studies
| Reagent / Tool | Function | Application Context |
|---|---|---|
| Error-Prone PCR Kit | Provides optimized mix of low-fidelity polymerase, biased dNTPs, and Mn2+ for efficient random mutagenesis [88]. | Directed Evolution: Library generation. |
| DNaseI | Enzyme used to randomly fragment genes for recombination-based DNA shuffling [88]. | Directed Evolution: Library generation. |
| NNK Degenerate Codons | Oligonucleotides containing these codons allow for saturation mutagenesis at targeted positions, creating all 19 amino acid variants [88]. | Directed & Semi-Rational Evolution: Focused library generation. |
| Biolog EcoPlates | Microplates pre-loaded with diverse carbon sources to profile the metabolic capabilities (traits) of microbial communities [76]. | Trait-Based Approaches: Community trait assessment. |
| Fluorescent Probes/Substrates | Colorimetric or fluorogenic enzyme substrates that enable high-throughput screening in microplates or via FACS [3] [88]. | Directed Evolution: Variant screening. |
| Phage or Yeast Display System | A platform for displaying protein variants on the surface of viruses or cells, allowing for affinity-based selection from highly complex libraries [88]. | Directed Evolution: Selection-based engineering (e.g., for antibodies). |
Integrated Workflows and Synthesis
The most powerful modern applications involve the sequential or synergistic integration of both paradigms. A common integrated workflow is the semi-rational directed evolution, depicted below.
Diagram Title: A Semi-Rational Integrated Workflow
This hybrid approach uses trait-based knowledge (e.g., from a crystal structure or AlphaFold model, phylogenetic analysis, or initial random mutagenesis data) to identify "hotspot" residues for targeted randomization [105] [88]. This dramatically reduces library size from the millions-billions required for fully random libraries to the thousands, making it feasible to screen the entire sequence space at those positions. The result is a more efficient search of the fitness landscape, accelerating the discovery of optimal variants.
The synergy extends to community engineering. Insights from trait-based studies of natural communities can identify which member species or functions are most critical to optimize. These specific functions can then be enhanced via directed evolution of the relevant enzymes in a single host, before reintroducing the optimized strain into a trait-designed consortium [9]. This combines the stability and emergent functionality of communities with the high-powered catalytic performance of evolved enzymes.
The quest to predict and engineer evolutionary pathways represents a frontier in biological research, with profound implications for drug development, synthetic biology, and understanding complex biological systems. Currently, two distinct computational approaches have emerged: trait-based evolution and directed evolution. Trait-based approaches focus on predicting evolutionary outcomes by modeling the complex interplay of phenotypic traits and environmental pressures, often using deep learning to navigate high-dimensional fitness landscapes. In contrast, directed evolution methodologies leverage machine learning to accelerate and guide the traditional "design-build-test-learn" cycle, actively steering evolutionary processes toward desired outcomes [23]. This article provides a comparative analysis of these paradigms, evaluating their experimental performance, methodological frameworks, and applicability to community optimization research.
Comparative Analysis of Evolutionary AI Approaches
Table 1: Comparison of Machine Learning Approaches for Evolutionary Pathway Prediction
| Feature | Trait-Based Evolution | AI-Directed Evolution |
|---|---|---|
| Theoretical Foundation | Models fitness landscapes from trait-environment interactions [107] | Engineering design as evolutionary process [23] |
| Primary Objective | Predict natural evolutionary trajectories & community dynamics | Engineer biological systems with customized functions [97] |
| Key Strengths | Captures emergent system behaviors; Models complex multi-species interactions | High throughput; Rapid optimization; Practical applications [108] |
| Data Requirements | Multi-omic datasets; Environmental parameters; Trait measurements | Targeted libraries; Performance metrics; Structural data |
| Typical Output | Predictive models of evolutionary dynamics | Novel biomolecules with engineered functions [108] [97] |
| Experimental Validation | Population dynamics monitoring; Fitness measurements | High-throughput screening; Functional characterization [108] |
| Implementation Scale | Communities, populations | Molecules, pathways, single organisms |
Table 2: Performance Metrics of Representative AI Systems in Evolutionary Design
| System/Platform | Approach Type | Key Achievement | Experimental Validation | Throughput/Scale |
|---|---|---|---|---|
| CRESt (MIT) [108] | AI-Directed Evolution | Discovered 8-element fuel cell catalyst with 9.3-fold improvement in power density per dollar over palladium | 3,500 electrochemical tests across 900 chemistries over 3 months | Robotic high-throughput synthesis & testing |
| AI-Driven De Novo Protein Design [97] | Trait-Based & Directed Hybrid | Creation of novel protein folds and functions not observed in nature | Experimental characterization of stability and function | Computational exploration of vast sequence-space |
| RoseTTAFold/AlphaFold | Trait-Based Prediction | Near-experimental accuracy in protein structure prediction | CASP competition validation; Crystallographic confirmation | Proteome-scale structure databases [97] |
Experimental Protocols and Methodologies
Protocol for AI-Directed Evolution (CRESt Platform)
The CRESt (Copilot for Real-world Experimental Scientists) platform exemplifies the modern AI-directed evolution workflow, which integrates robotic equipment and multimodal learning [108]:
- Knowledge Integration: The system begins by creating embeddings of potential material recipes using previous literature text and databases before conducting experiments.
- Search Space Reduction: Principal component analysis is performed in this knowledge embedding space to derive a reduced search space capturing most performance variability.
- Experimental Design: Bayesian optimization is applied in the reduced space to design new experiments.
- Robotic Execution: Liquid-handling robots and carbothermal shock systems synthesize candidate materials based on optimized recipes.
- High-Throughput Characterization: Automated electron microscopy, optical microscopy, and electrochemical workstations characterize synthesized materials.
- Multimodal Learning: Newly acquired experimental data and human feedback are incorporated into a large language model to augment the knowledge base and redefine the search space.
- Iterative Optimization: The process repeats with continuous refinement of candidate materials through multiple generations.
This methodology enabled CRESt to explore over 900 chemistries and conduct 3,500 electrochemical tests, leading to the discovery of a catalyst material that delivered record power density in a fuel cell [108].
Protocol for AI-Driven Protein Design
AI-driven de novo protein design represents a fusion of trait-based prediction and directed evolution principles [97]:
- Functional Specification: Define desired protein function, stability parameters, and structural constraints.
- Generative Design: Use deep learning models (e.g., RoseTTAFold, AlphaFold) to generate amino acid sequences predicted to fold into structures capable of the target function.
- In Silico Screening: Computational models predict protein stability, folding kinetics, and functional properties for thousands of candidate sequences.
- Library Construction: Synthesize genes encoding promising candidate proteins.
- Experimental Characterization: Express and purify proteins for functional assays and structural validation (e.g., X-ray crystallography, NMR).
- Iterative Refinement: Use experimental data to retrain predictive models and generate improved designs.
This approach has successfully created novel enzymes, protein-based therapeutics, and biomaterials with functions not found in nature [97].
Visualization of Methodological Frameworks
The Evolutionary Design Spectrum
AI-Directed Evolutionary Workflow (CRESt)
The Scientist's Toolkit: Essential Research Reagents and Platforms
Table 3: Essential Research Reagents and Platforms for AI-Driven Evolution
| Reagent/Platform | Function | Application Context |
|---|---|---|
| CRESt Platform [108] | Multimodal AI system for materials discovery | Integrates literature knowledge, robotic experimentation, and active learning for accelerated materials optimization |
| RoseTTAFold/AlphaFold [97] | Protein structure prediction & design | De novo protein design and functional site engineering |
| Liquid-Handling Robots [108] | Automated sample preparation and synthesis | Enables high-throughput experimentation for library screening |
| Carbothermal Shock System [108] | Rapid synthesis of materials | Quick generation of material variants for testing |
| Automated Electrochemical Workstation [108] | High-throughput functional testing | Rapid performance characterization of catalyst materials |
| Generative AI Models (VAEs, GANs) | Novel molecular structure generation | Exploration of chemical space for drug discovery and protein design |
| Feast/Tecton Feature Stores | Real-time feature management for ML | Streaming data infrastructure for continuous model training |
Discussion: Integration Frontiers and Future Directions
The comparative analysis reveals that trait-based and directed evolution approaches, while distinct in methodology, are increasingly converging in modern biological research. Trait-based models provide the foundational understanding of evolutionary constraints and possibilities, while directed evolution offers a practical engineering framework for achieving targeted outcomes. The most significant advances are emerging from hybrid approaches that leverage the predictive power of trait-based modeling to inform and accelerate directed evolution campaigns [97] [23].
Future developments will likely focus on several key areas: (1) improved integration of multiscale biological data (genomic, proteomic, metabolic) to create more accurate trait-based predictors; (2) enhanced closed-loop AI systems that tightly couple prediction, design, and experimental validation with minimal human intervention; and (3) expansion of these approaches to complex community-level optimization, where multiple organisms and their interactions must be considered simultaneously [108] [97] [23].
For drug development professionals, these advances translate to dramatically accelerated discovery timelines and access to novel biological space previously inaccessible through conventional methods. The successful application of these technologies to problems like fuel cell catalyst discovery and de novo protein design demonstrates their readiness for addressing critical challenges in therapeutic development and sustainable biotechnology [108] [97].
Conclusion
The strategic choice between trait-based and directed evolution approaches is not a binary one but a question of context and goal. Trait-based ecology provides a powerful lens for understanding and predicting the performance of natural communities, while directed evolution offers an unparalleled ability to create novel biological functions in the laboratory. The key takeaway is that these approaches are increasingly synergistic. Insights from trait-based studies can inform the design of smarter directed evolution libraries, and the principles of iterative selection can refine our understanding of trait-function relationships. For the future of biomedical and clinical research, this integration promises a new generation of optimized cell lines for bioprocessing, engineered microbial consortia for therapeutic purposes, and novel biocatalysts for drug synthesis, all achieved with greater speed, yield, and predictability. Embracing this combined framework will be crucial for tackling complex challenges in drug development and synthetic biology.
References
- 1. Rapid evolution of quantitative traits: theoretical perspectives [https://pmc.ncbi.nlm.nih.gov/articles/PMC3894905/]
- 2. The evolution of trait variance creates a tension between ... [https://www.nature.com/articles/s41467-022-30090-4]
- 3. A primer to directed evolution: current methodologies and ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10074555/]
- 4. Directed Evolution: Past, Present and Future - PMC [https://pmc.ncbi.nlm.nih.gov/articles/PMC4344831/]
- 5. Navigating directed evolution efficiently: optimizing ... [https://www.frontiersin.org/journals/molecular-biosciences/articles/10.3389/fmolb.2024.1439259/full]
- 6. Directed evolution: new parts and optimized function - PMC [https://pmc.ncbi.nlm.nih.gov/articles/PMC2775421/]
- 7. Long-term studies provide unique insights into evolution - PMC [https://pmc.ncbi.nlm.nih.gov/articles/PMC12359119/]
- 8. Trait trajectories in evolving populations: insights from mathem... [https://evolbiol.peercommunityin.org/articles/rec?id=555]
- 9. Towards synthetic ecology: strategies for the optimization ... [https://www.frontiersin.org/journals/synthetic-biology/articles/10.3389/fsybi.2025.1532846/full]
- 10. Trait-based modelling in ecology: A review of two decades ... [https://www.sciencedirect.com/science/article/abs/pii/S0304380019301735]
- 11. Directed Evolution of Microbial Communities in Fermented ... [https://www.mdpi.com/2304-8158/14/2/216]
- 12. Trait-based ecology, trait-free ecology, and in between [https://pubmed.ncbi.nlm.nih.gov/39410833/]
- 13. Trait-based community assembly and succession of the ... [https://www.nature.com/articles/s41467-019-08377-w]
- 14. Trait-based community assembly and functional strategies ... [https://www.frontiersin.org/journals/plant-science/articles/10.3389/fpls.2024.1451981/full]
- 15. Synthetic directed evolution for targeted engineering of plant ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC11402689/]
- 16. Ecological design of high-performance synthetic microbial ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12373479/]
- 17. A chimeric viral platform for directed evolution in ... [https://www.nature.com/articles/s41467-025-59438-2]
- 18. Directed evolution [https://en.wikipedia.org/wiki/Directed_evolution]
- 19. Spiegelman's Monster: a milestone in molecular biology [https://oligofastx.com/spiegelmans-monster-a-milestone-in-molecular-biology/]
- 20. Optimisation strategies for directed evolution without ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC11698521/]
- 21. Protein Engineering and Drug Discovery [https://www.mdpi.com/2218-273X/15/11/1628]
- 22. Generative AI for drug discovery and protein design [https://www.sciencedirect.com/science/article/pii/S2590098625000107]
- 23. Engineering is evolution: a perspective on design ... [https://www.nature.com/articles/s41467-024-48000-1]
- 24. Functional traits, the phylogeny of function, and ecosystem ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC3790543/]
- 25. Co-Selection of Resistance to Antibiotics, Biocides and Heavy ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC4790313/]
- 26. Co-selection for antibiotic resistance by environmental ... [https://www.nature.com/articles/s44259-024-00026-7]
- 27. The sequence-structure relationship and protein function ... [https://pubmed.ncbi.nlm.nih.gov/19406632/]
- 28. Machine Learning for Protein Engineering - PMC [https://pmc.ncbi.nlm.nih.gov/articles/PMC10246115/]
- 29. The simplicity of protein sequence-function relationships [https://www.nature.com/articles/s41467-024-51895-5]
- 30. Current successes and remaining challenges in protein ... [https://www.frontiersin.org/journals/bioinformatics/articles/10.3389/fbinf.2023.1222182/full]
- 31. Synthetic directed evolution in plants: unlocking trait ... - PMC [https://pmc.ncbi.nlm.nih.gov/articles/PMC8434914/]
- 32. Synthetic directed evolution for targeted engineering of ... [https://www.frontiersin.org/journals/plant-science/articles/10.3389/fpls.2024.1449579/full]
- 33. Inferring protein sequence-function relationships with large ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC7856229/]
- 34. Random Mutagenesis: 8 Approaches for Success [https://bitesizebio.com/252/8-approaches-to-random-mutagenesis/]
- 35. DNA shuffling [https://en.wikipedia.org/wiki/DNA_shuffling]
- 36. Fueling chromosomal gene diversification and artificial ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12455791/]
- 37. Advancements in CRISPR/Cas systems for disease treatment [https://pmc.ncbi.nlm.nih.gov/articles/PMC12254762/]
- 38. The hidden risks of CRISPR/Cas: structural variations and ... [https://www.nature.com/articles/s41467-025-62606-z]
- 39. DNA Shuffling | MyBioSource Learning Center [https://www.mybiosource.com/learn/testing-procedures/dna-shuffling/]
- 40. Beyond directed evolution - semi-rational protein ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC2982887/]
- 41. Beyond the Natural Proteome: Nondegenerate Saturation ... [https://www.sciencedirect.com/science/article/abs/pii/S0076687916303688]
- 42. Random Mutagenesis as a Promising Tool for Microalgal ... [https://www.mdpi.com/1660-3397/20/7/440]
- 43. Saturation mutagenesis [https://en.wikipedia.org/wiki/Saturation_mutagenesis]
- 44. Using Site-Saturation Mutagenesis to Explore Mechanism ... [https://pubmed.ncbi.nlm.nih.gov/23895593/]
- 45. Site-saturation mutagenesis of 500 human protein domains [https://www.nature.com/articles/s41586-024-08370-4]
- 46. Boosting the efficiency of site-saturation mutagenesis for a ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC6013526/]
- 47. Optimisation strategies for directed evolution without sequencing [https://journals.plos.org/ploscompbiol/article?id=10.1371/journal.pcbi.1012695]
- 48. BD FACSAriaâ„¢ Fusion Flow Cytometers [https://www.bdbiosciences.com/en-us/products/instruments/flow-cytometers/research-cell-sorters/bd-facsaria-fusion]
- 49. BD FACSAriaâ„¢ III | High Sensitivity Flow Cytometer [https://www.bdbiosciences.com/en-us/products/instruments/flow-cytometers/research-cell-sorters/bd-facsaria-iii]
- 50. Advancements in research leveraging phage display ... [https://www.frontiersin.org/journals/microbiology/articles/10.3389/fmicb.2025.1632560/full]
- 51. Utilizing a high-throughput visualization screening ... [https://www.sciencedirect.com/science/article/abs/pii/S0956566324008121]
- 52. BD FACSAria Fusion | Faculty of Natural Sciences [https://www.imperial.ac.uk/natural-sciences/departments/life-sciences/research/flow-cytometry-facility/equipment/fusion/]
- 53. BD FACSCaliburâ„¢ Flow Cytometer [https://www.bdbiosciences.com/en-nl/products/instruments/flow-cytometers/clinical-cell-analyzers/facscalibur]
- 54. High-throughput proteome integral solubility alteration ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12474936/]
- 55. Biocatalyst Development by Directed Evolution - PMC [https://pmc.ncbi.nlm.nih.gov/articles/PMC3351540/]
- 56. Enzymes as Catalysts in Industrial Biocatalysis: Advances ... [https://www.mdpi.com/2073-4344/15/9/891]
- 57. Understanding activity-stability tradeoffs in biocatalysts by ... [https://www.nature.com/articles/s41467-024-45630-3]
- 58. Considerations when Measuring Biocatalyst Performance [https://pmc.ncbi.nlm.nih.gov/articles/PMC6804192/]
- 59. Reflections from Biotrans 2025: Charting the Future of… [https://www.biocatalysts.com/media-resources/biotrans-biocatalysis-future-trends]
- 60. Automated in vivo enzyme engineering accelerates ... [https://www.nature.com/articles/s41467-024-46574-4]
- 61. Machine learning applied to biocatalysis research [https://www.nature.com/articles/s41467-025-64510-y]
- 62. Protein Optimization 101: Insights from the literature [https://www.adaptyvbio.com/blog/po101/]
- 63. Data-Driven Yield Improvement in Upstream Bioprocessing ... [https://www.mdpi.com/2227-9717/13/11/3394]
- 64. Bioprocess Optimization - Eppendorf Middle East & Africa [https://www.eppendorf.com/us-en/lab-academy/applied-industries/bioprocessing/production-of-antibodies-and-hormones/bioprocess-optimization/]
- 65. Bioprocess Yield Troubleshooting: Effective Strategies [https://www.linkedin.com/advice/1/your-bioprocess-optimization-faces-unexpected-yield-oiqze]
- 66. Dynamic nonlinear modeling improves product-to-biomass ... [https://www.sciencedirect.com/science/article/pii/S2001037024003131]
- 67. Active learning-assisted directed evolution [https://www.nature.com/articles/s41467-025-55987-8]
- 68. Optimal adaptive control of a bioprocess with yield– ... [https://www.sciencedirect.com/science/article/abs/pii/S0168165698001229]
- 69. Decadal decline of functional diversity despite increasing ... [https://www.sciencedirect.com/science/article/pii/S1470160X24006009]
- 70. Unraveling the seasonality of functional diversity through ... [https://www.nature.com/articles/s43247-025-02646-x]
- 71. Functional diversity buffers the effects of a pulse ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC8601937/]
- 72. Impacts of Urban Areas and Their Characteristics on Avian ... [https://www.frontiersin.org/journals/ecology-and-evolution/articles/10.3389/fevo.2017.00084/full]
- 73. Patterns of Functional Diversity, Species ... - PubMed Central [https://pmc.ncbi.nlm.nih.gov/articles/PMC12058463/]
- 74. TbasCO: trait-based comparative 'omics identifies ... [https://www.nature.com/articles/s43705-022-00189-2]
- 75. Rethinking Ecological Measures Of Functional Diversity [https://arxiv.org/html/2506.17839v1]
- 76. Trait-based approaches for understanding microbial ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC4033906/]
- 77. Aspects and Implementation of Pharmaceutical Quality by ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12114689/]
- 78. PAT implementation for advanced process control in solid ... [https://www.sciencedirect.com/science/article/abs/pii/S037851732101214X]
- 79. Advanced control system for upstream biomanufacturing ... [https://www.pharmafocusamerica.com/manufacturing/advanced-model-predictive-control-system]
- 80. Trends and new process analytical technologies in ... [https://www.sciencedirect.com/science/article/abs/pii/S037851732500794X]
- 81. Automating Personal Workflows with MCP | by Pratham Sharma [https://mehmehsloth.medium.com/automating-personal-workflows-with-mcp-d1f3b9f7f26c]
- 82. Best MCP Automation Platforms for Enterprise [https://www.truefoundry.com/blog/mcp-automation-platforms-for-enterprise]
- 83. Continuous-Trait Probabilistic Model for Comparing Multi- ... [https://www.sciencedirect.com/science/article/pii/S2405471218302400]
- 84. How Size Matters: Diversity for Fragment Library Design [https://pmc.ncbi.nlm.nih.gov/articles/PMC6696339/]
- 85. New directions in library design and analysis [https://www.sciencedirect.com/science/article/abs/pii/S1367593108000380]
- 86. Methods for community-engaged data collection and ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC11978136/]
- 87. Bayesian optimization with evolutionary and structure-based ... [https://almob.biomedcentral.com/articles/10.1186/s13015-021-00195-4]
- 88. A Technical Guide to Directed Evolution for Enhancing ... [https://www.ranomics.com/a-technical-guide-to-directed-evolution-for-enhancing-protein-stability-and-function]
- 89. Fitness Costs and the Rapid Spread of kelch13-C580Y ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC6125530/]
- 90. Fitness effects of CRISPR endonucleases in Drosophila ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC9545523/]
- 91. CRISPR-enabled investigation of fitness costs associated ... [https://www.frontiersin.org/journals/plant-science/articles/10.3389/fpls.2023.1278133/full]
- 92. COmmunity and Single Microbe Optimisation System ... [https://www.nature.com/articles/s41540-025-00534-w]
- 93. Using Directed Evolution to Develop New Vectors ... [https://globalgenes.org/raredaily/using-directed-evolution-to-develop-new-vectors-for-genetic-medicines/]
- 94. The optimization of microbial functions through rational ... [https://pubmed.ncbi.nlm.nih.gov/38372207/]
- 95. Microbial functional diversity: From concepts to applications [https://pmc.ncbi.nlm.nih.gov/articles/PMC6822047/]
- 96. Functional Diversity - an overview [https://www.sciencedirect.com/topics/agricultural-and-biological-sciences/functional-diversity]
- 97. The Role of AI-Driven De Novo Protein Design in the ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12467925/]
- 98. The evolution of trait variance creates a tension between ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC9085882/]
- 99. Semantic design of functional de novo genes from a ... [https://www.nature.com/articles/s41586-025-09749-7]
- 100. Comparative analysis, diversification, and functional ... [https://www.nature.com/articles/s41598-024-62876-5]
- 101. Recent applications, future perspectives, and limitations of ... [https://www.sciencedirect.com/science/article/pii/S216225312500188X]
- 102. Challenges and Opportunities in the Application of CRISPR ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12503735/]
- 103. De-extinction and beyond: trait design powered by ... [https://www.sciencedirect.com/science/article/abs/pii/S0167779925004032]
- 104. Design of highly functional genome editors by modelling ... [https://www.nature.com/articles/s41586-025-09298-z]
- 105. Directed evolution of hydrocarbon-producing enzymes [https://biotechnologyforbiofuels.biomedcentral.com/articles/10.1186/s13068-025-02689-4]
- 106. Directed Evolution for the Discovery of Engineered ... [https://pubmed.ncbi.nlm.nih.gov/40660468/]
- 107. On the Parallels Between Evolutionary Theory and ... [https://arxiv.org/html/2505.23774v1]
- 108. AI system learns from many types of scientific information ... [https://news.mit.edu/2025/ai-system-learns-many-types-scientific-information-and-runs-experiments-discovering-new-materials-0925]